11 a sejtek evolúciója 1. a molekuláktól az első sejtekig (prebiológiai evolúció) egyszerű biomolekulák prebiotikus körülmények közö
11
A sejtek evolúciója
1. A molekuláktól az első sejtekig (prebiológiai evolúció)
Egyszerű biomolekulák prebiotikus körülmények között is képződtek
Földünk kb. 4.6 milliárd éve keletkezett. A Föld első, hosszabb ideig
megmaradó légköre redukáló jellegű volt, és bizonyos, hogy sem
oxigént, sem ózont nem tartalmazott. Ennek az volt a következménye,
hogy a világűrből érkező különféle sugárzások szabadon eljuthattak a
felszínre. Haldane és Oparin az 1920-as években vetették fel az élet
kialakulásának spontán lehetőségét ilyen oxigénmentes környezetben.
Urey professzor (a Chicagói Egyetem oktatója) könyvet irt az
égitestekről “The Planets” címmel, melyben megadta a Föld
feltételezett légkörét. A professzor Miller nevű tanítványa pedig
megpróbálta laboratóriumi kísérletben ellenőrizni, hogy milyen
egyszerű szerves vegyületek képződhettek ebben a légkörben az
Oparin-Haldane hipotézis szerint. Miller gázkeveréket (CH4, NH3 és H2)
melegített vízzel, és elektromos szikrakisüléseket hozott létre,
miközben hűtőn kondenzáltatta a vízgőzt és visszavezette azt az
edénybe (ld. 1. ábra).
1. ábra: Az (Ure -) Miller-féle kísérlet
2. ábra: Néhány egyszerűbb vegyület, melyek az Urey - Miller
kísérletben képződtek
A vízgőz és a gázelegy alkotórészei az elektromos kisülések aktiváló
hatására reakcióba lépetek egymással, és két hét után a 2. ábrán
bemutatott vegyületeket lehetett kimutatni az oldatból. Természetesen
a valóságban sokkal több idő (több 100 millió év) állt rendelkezésre a
szerves vegyületek abiogén kialakulására.
A felsorolt vegyületek további reakciókba léptek egymással illetve
más, az ősóceánban megtalálható szervetlen vegyületekkel. A
hidrogén-cianidból (HCN) pl. nukleinsavbázisok (pl. adenin – (HCN)5 )
keletkeztek, míg a formaldehidből (CH2O) szénhidrátok (pl. ribóz – (CH2O)5
) képződtek.
Végeredményben elmondható, hogy a mai élőlényekben előforduló szerves
molekulafajták minden reprezentánsa keletkezett az Urey-Miller
kísérletben: karbonsavak, aminosavak, cukrok, purin és pirimidin
bázisok. Ezek az anyagok építőkövei a mai élőlényekben előforduló
makromolekuláknak mint pl. az RNS, a DNS és a fehérjék. Tulajdonképpen
sokkal több képviselője keletkezett ezeknek az építőköveknek, mint
amit mai élőlények használnak. A ma élő sejtek fehérjéiben csak 20
féle aminosav, a nukleinsavaikban pedig csak 5 féle nukleotid fordul
elő.
Az Urey-Miller kísérletben az aminosavaknak és szénhidrátoknak két
optikailag eltérő módosulata keletkezett, melyek a polarizált fényt
ellentétes irányba forgatják (D és L konfigurácó). A mai élőlényekben
viszont az aminosavaknak csak az L-formája (kivéve: baktériumok
sejtfalát) és a szénhidrátoknak a D-formája fordul elő. Ennek a
jelenségnek nem ismert még a magyarázata.
Az abiogén úton keletkező szerves vegyületek az ősi Föld körülményei
között az ősóceánban oldott formában lehettek jelen. Ebben az oldatban
– a párolgás hatására – több helyen szerves vegyületek nagy
koncentrációja alakulhatott ki, ami további reakciókat tett lehetővé.
Az aminosavak és nukleotidok spontán polimerizálnak
A
z aminosavak peptidkötés létesítésével, a nukleotidok pedig
foszfodiészter kötés képződése mellett polimerizálódhatnak. Mindkét
reakció vízképződéssel jár, ezért híg vizes oldatban nem
kedvezményezettek, azaz a polimerizációhoz energiára van szükség. Ezt
a problémát a mai élőlények úgy oldják meg, hogy aktivált
építőelemeket polimerizálnak (s így nem víz, hanem foszforsav lép ki
az összekapcsolódás során). Hogyan játszódhatott le a polimerizáció
régen, az élet kialakulása előtt?
3. ábra: Aminosavak és nukleotidok spontán polimerizációja.
Az ősi Földön a polimerek kialakulásának az alábbi lehetőségei
jöhettek számításba:
– Nem vizes közegben történő polimerizáció az ősóceán beszáradó
részein. Ennek bizonyítékául Sidney Fox kísérleteiben az aminosavak
por alakú keverékénél 130-150oC-ra történő hevítést követően
polipeptideket mutatott ki.
– Bernal elmélete szerint vizes közegben agyagásványok katalizálhatták
az első polimerizációkat, vagyis az ősóceán partján, a „pocsolyákban”
lévő töményebb oldatokban keletkezhettek az első biopolimerek.
Az élőlények által szintetizált biopolimerekkel szemben azonban
ezekben az abiotikus úton végbemenő polimerizációkban a nukleinsavak
és a fehérjék változó lánchosszúságban és véletlenszerű összetétellel
keletkeztek.
A sejtek alapvető tulajdonsága az önreprodukciós képesség, vagyis hogy
önmagukkal azonos utódot képesek létrehozni (egy sejtből két sejt
lesz). Kémiai rendszerekben sem ismeretlen ez a képesség, de a
kémiában inkább autokatalitikusnak hívjuk azt a folyamatot, amikor a
reakció végterméke elősegíti önmaga képződését. Fontos kérdés, hogy a
önreprodukciós képesség mikor jelent meg az evolúció során?
Az autokatalízis megjelenését Gánti Tibor chemoton elmélete
membránokba zárt a kémiai körfolyamatokban, míg Manfred Eigen
hiperciklus elmélete a makromolekulák képződésében jelöli meg
elsőként.
4. ábra A polinukleotidok saját szintézisüket képesek irányítani
Melyik makromolekulának van autokatalitikus tulajdonsága? A
polipeptidek jó katalizátorok, de nem tudják saját szintézisüket
nukleinsavak nélkül irányítani. Ezzel szemben a polinukleotidok
képesek önmaguk szintézisének irányítására komplement pár képzés révén
(ld. 4. ábra).
5. ábra: Két másolás után visszakpajuk az eredeti polinukleotidot.
Az egyik szál templátként szolgál egy másik szál képződéséhez. Például
egy poli-C egy poli G lánc felépülését irányíthatja és vice versa.
Könnyű belátni (5. ábra), hogy két másolás után visszakapjuk az
eredeti polinukleotid láncot (templát -> replika -> templát).
Ez a mechanizmus azonban csak a potenciális lehetőségét teremti meg az
önreprodukciós képességnek, mert a folyamat önmagában nagyon lassú, és
ahhoz, hogy mérhető sebességgel játszódjon le, valamilyen formában
katalízisre van szükség. Manapság a katalitikus funkciót a specifikus
fehérjék, enzimek biztosítják a nukleinsavak másolásához. Ha
végiggondoljuk a dolgot, máris a legősibb tyúk-tojás problémával
találjuk magunkat szemben: mi volt előbb? Nukleinsavak vagy fehérjék?
A mai élőlényekben az RNS-ek szintézisét fehérjék katalizálják, az
RNS-ek pedig a fehérjeszintézist segítik. A prebiotikus (ős-) leves-ben
lehettek primitív polipeptidek, de az RNS molekulák ritkák maradtak,
ha ők maguk nem tudták a saját szintézisüket serkenteni. Sokáig azt
hittük, hogy a nukleinsavak csak információ tárolásra és átvitelre
képesek, de katalízisre nem.
Specializált RNS-ek biokémiai reakciókat katalizálnak
Az önreprodukáló struktúrák létrejöttének megértésében fontos szerepe
volt annak a néhány évvel ezelőtti felismerésnek, miszerint az RNS-ek
kémiai reakciók katalízisére is képesek. A katalitikus RNS neve
ribozim.
Az RNS katalitikus tulajdonságát háromdimenziós szerkezete
(feltekeredése) határozza meg. Az RNS molekula feltekeredési módja
pedig a szekvenciájából következik, a molekulán belüli komplementer
(kiegészítő) szakaszok ugyanis párosodni tudnak egymással: pl. egy
GGGG párosodni tud egy CCCC-vel azonos molekulán belül és egy komplex
3 dimenziós szerkezet jön létre.
Az önreprodukáló molekulák természetes szelekciója
Tételezzük fel, hogy létrejött egy megfelelő nukleotid sorrendű RNS,
mely katalizátora lehetett egy másik RNS templát polimerizációs
replikációjának. Abban az esetben, ha ez a templát éppen a saját
kópiája, ez a bizonyos RNS molekula autokatalitikusan elszaporodik.
Elvileg több ilyen autokatalitikus rendszer is kialakulhatott, és
ekkor megindult köztük a versengés. Miért versenyeztek? A versengés az
építőkövekért, vagyis a táplálékért folyt. A nukleinsavak másolása
során hibák is keletkezhettek, és az új kópiák nem feltétlenül voltak
azonosak a kiindulási templáttal. Könnyen észrevehetjük, hogy azok a
változatok szaporodtak el az építőkövekért folytatott versenyben,
melyek katalitikus aktivitásukat javítani tudták a “szüleikhez” képest
vagyis rátermettebbek lettek. Megfigyelhetjük, hogy az önreprodukáló
molekulák körében a természetes szelekció (variáció és a rátermettebb
elszaporodása) érvényesül.
Információ áramlás a polinukleotidoktól a polipeptidekig
3.5 - 4 milliárd évvel ezelőtt a Földön valószínűleg önreprodukáló RNS
molekulák és különféle polipeptidek lehettek jelen. A makromolekulák
között ugyanolyan verseny folyt az prekurzorokért, akárcsak mai a
élőlények közötta táplálékért. De ki nyerte a végső versenyt? Hosszú
távon semmiképpen nem azok az RNS-ek, akik maximálták a katalitikus
aktivitásukat. Az RNS-ek katalitikus aktivitása ugyanis korlátozott!
Hosszú távon az az autokatalitikus rendszer nyerte a versenyt,
amelyben a katalitikus RNS-ek önzetlen alapon segítették olyan RNS-ek
másolását, amelyeknek nem volt katalitikus funkciója az RNS-ek
másolásában. Ezek az RNS-ek így más funkcióra specializálódhattak és
ez a más funkció a polipeptidek szintézisének irányítása volt.
De hogyan tudja egy polinukleotid egy teljesen másfajta polimer
(fehérje) aminosav sorrendjét meghatározni? Ehhez a következőkre van
szükség:
- polipeptid sorrendjét meghatározó RNS templátra,
- az aminosavakat polipeptiddé kapcsoló katalizátor RNS-re, és
- olyan adapter RNS molekulákra, amelyek specifikus aminosavakat
kötnek.
A templát és az adapter molekulák a komplementer bázispár képzés elve
alapján összekapcsolódnak, és ezáltal a templát az adaptereken
keresztül meghatározza a fehérje aminosav sorrendjét. A katalizátor
RNS-ek pedig összekapcsolták az aminosavakat, és így mehetett végbe az
ősi protein-szintézis.
Ma a protein szintézist a riboszómák végzik, melyek RNS-ekből
(rRNS-ek) és fehérjékből álló szerkezetek. Az a tény, hogy a mai
fehérjeszintézisben is fontos szerepe van az rRNS-eknek, sejtetni
engedi, hogy az ősidőkben egyedül ők voltak a katalizátorok.
Az RNS irányította protein szintézishez szükség van egy kódra, aminek
alapján a nukleinsav szekvencia meghatározza a fehérje aminosav
sorrendjét. Ez a kód látszólag teljesen azonos a ma élő élőlényekben,
ezért feltehetően azok mind egy közös őstől erednek.
Ez a közös ős az RNS-irányított proteinszintézis kimunkálásával olyan
fehérjék szintézisét oldotta meg, melyek hasznos gépeknek
bizonyulhattak:
- az RNS-ek másolásában (pl. RNS polimeráz), valamint
- a gépek (proteinek) gyártásában (riboszóma fehérjék).
Mivel a fehérjék sokkal jobb katalizátorok, mint a nukleinsavak, ezért
ez az ős olyan előnyhöz jutott az evolúciós küzdelemben, hogy ezt a
vívmányát sohasem feledte el.
Membrán definiálta az első sejtet
Az RNS segítségével szintetizálódott fehérjemolekula nem feltétlenül
csak azt a bizonyos RNS-t tudta segíteni a reprodukciójában, ami őt
létrehozta, amíg ki nem alakult a plazmamembrán. A plazmamembrán
kialakulása egy meghatározott kompartment létrehozását jelenti,
amelyben együtt maradhatnak azok az RNS-ek és fehérjék, melyek egymás
képződését segítetik elő. Ez a funkció egy másik molekula csoport
által valósulhatott meg, amely alkalmasnak bizonyult határfelület
létrehozására. A hidrofil ás hidrofób részekből álló foszfolipidek
kiválóan alkalmasak erre a funkcióra, mivel vizes oldatban spontán
kettős réteget (bilayer) alkotnak (6. ábra). A prebiotikus levesben
tehát a spontán aggregálódó foszfolipidek vehették körül a replikálódó
RNS és fehérjemolekulákat.
6. ábra: A foszfolipidek egyszeres vagy kétszeres réteget
alkotnak.
Minden ma élő sejtben a DNS az örökítő anyag
Idáig egy spekulatív prebiológiai történettel ismerkedtünk meg, amely
talán fő vonalaiban érvényes, és aminek eredményeként 3-4 milliárd
évvel ezelőtt kialakult az első primitív sejt. Ez a sejt feltehetően a
ma élő legegyszerűbb és legkisebb (átmérőjük 0.3 µm) sejtekhez, az ún.
mikoplazmákhoz (csupasz, sejtfal nélküli baktériumok) volt hasonló.
Van azonban egy döntő különbség a Földön kialakult első sejtek és a ma
élő legegyszerűbb sejtek között: nevezetesen, hogy a mai “modern”
sejtekben kivétel nélkül a DNS az örökítő anyag és nem az RNS. Ez azt
jelenti, hogy az evolúció során valamikor az RNS molekulák, melyek
kezdetben mind információ tároló és katalitikus szerepet is
betöltöttek, nemcsak a katalitikus funkciójukat adták át a
fehérjéknek, hanem az információt hordozó szerepüket is elvesztették
illetve átadták a DNS-nek. Ezt követően az RNS-ek mint összekötő
kapocs maradtak meg a DNS és a fehérjék között (7. ábra).
A prokarióták kialakulása és fejlődése
A ma élő sejtek közös őse tehát mintegy 3-4 milliárd évvel ezelőtt
születhetett meg, és kedvező tulajdonságainak köszönhetően
túlszaporodott a vetélytársain. Ennek a sejtnek feltehetően nem volt
különösebb belső struktúrája, és ma élő legközelebbi rokonait a Földön
prokarióta (elősejtmagvas: pro = előtt, karion = mag) sejteknek
hívjuk.
A prokarióták (baktériumok) tehát a közös ős legközelebbi rokonai,
akik 2 milliárd éven keresztül egyedül uralták a Földet. Ezek ma is a
legegyszerűbb organizmusok a Földön, amelyek igen változatos formákat
illetve alakot öltenek (ld. 8. ábra). Méretük mikrométer nagyságrendű.
Sejtfelépítésük azonban már sokkal komplikáltabb mint a közös ősé
volt, hiszen a mikoplazmákat leszámítva sejtmembránjukat sejtfal
határolja. A természetben a legváltozatosabb helyeken előfordulnak,
mert biokémiailag igen változatosak.
8. ábra: A prokarióták alakja és mérete igen változatos.
A prokarióták két nagy csoportját szokás megkülönböztetni:
- eubaktériumok (valódi baktériumok),
- ősbaktériumok (archaebacteria).
Az ősbaktériumok olyan szokatlan élőhelyeken élnek, mint mocsarak,
tengerek mélye vagy hőforrások, amelyekben a körülmények nagyon
hasonlítanak az ősi Földön uralkodó viszonyokhoz. Ebből régebben arra
következtettek, hogy ennek a prokarióta csoportnak a tagjai alakultak
ki először a Földön, és erre utal az ősbaktérium elnevezés is. A
molekuláris vizsgálatok azonban kiderítették, hogy ez az állítás nem
érvényes és az ősbaktériumok semmivel sem ősibbek a valódi
baktériumoknál. Mindössze arról van szó, hogy e két prokarióta csoport
fejlődése nagyon korán elvált egymástól.
Az ősbaktériumok három nagy csoportját szokás megkülönböztetni:
- termoacidofilek: forró, savas környezetben (pH = 2 és t=80oC) élnek
(pl. Thermoplasma acidophilum (“csodaszervezet”).
- extrém halofilek: szinte telitett sóoldatban élnek (pl.
Halobacterium).
- metanogének: a CO2 redukciójával metánt állítanak elő (metánképzők):
CO2 + 4 H2 ---> CH4 + 2 H2O
A mai baktériumok sejtfelszíne különböző
Amint azt említettük a mai baktériumok sejtfallal rendelkeznek, ami
lehet vastag vagy vékony szerkezet. Ennek eldöntésére szolgál az ún.
Gram festés, ami a kristályibolya-jód komplex alkoholos kezelés
hatására történő színeződést vizsgálja. A vastag sejtfallal rendelkező
őn. Gram pozitív batériumok festődnek, míg a sokkal vékonyabb sejtfalú
Gram negatívok nem. A Gram festésben mutatkozó eltérések a sejtfelszín
struktúrájában illetve azon belül is a sejtfal vastagságában
keresendők. Ennek illusztrálására a ábrán bemutatjuk a Gram pozitív és
Gram negatív valódi baktériumok sejtfelszínének jellegzetes
struktúráját. Láthatóan a Gram pozitív baktériumoknál a
citoplazmamembránon kívül mindössze a vastag sejtfal (murein) foglal
helyet. Ezzel szemben a Gram negatív baktériumoknál egy máshol elő nem
forduló struktúra, a külső membrán határolja a sejtet. A vékony
sejtfal ebben az esetben a két membrán között foglal helyet, az ún.
periplazmás térben.
Metabolikus reakciók fejlődése a bakteriális evolúció során
A mai baktériumokban enzimek százai energiát és építőköveket gyártanak
az ún. metabolikus útvonalakon. Régen azonban ezeknek az építőköveknek
és energiában gazdag molekuláknak a többsége szabadon rendelkezésre
állt a környezetben, az ősóceán tehát szinte maga a “Paradicsom” volt
az első sejtek számára. Mivel - mint már említettük- O2 nem volt jelen
a légkörben, ezért az első sejtek anaerobok (O2 hasznosítására
képtelenek) voltak, és szerves vegyületekre voltak utalva (heterotrófok
ill. organotrófok). Az általuk felhasznált szerves vegyületek
abiotikusan keletkeztek az ősi Földön. Az élet kialakulását követően
azonban ezen szerves molekulák mennyisége rohamosan csökkent a
környezetükben, mivel a sejtek folyamatosan felélték, felhasználták
azokat. A környezet tehát állandóan romlott a sejtek számára, ezért
azok a sejtek, amelyek alkalmazkodni tudtak az állandóan romló
feltételekhez, hatalmas evolúciós előnyre tettek szert a többiekkel
szemben. Ez az alkalmazkodás új molekulák és új energiaforrások
hasznosítását jelentette. Ehhez természetesen új enzimek kialakulására
volt szükség, minek következtében a sejtek enzimkészlete növekedett.
Ez az alkalmazkodás a változó környezetekhez azonban nem érintette a
metabolizmus alapreakcióit (“gyökerét). Ennek következtében ezek a
reakciók minden ma élő sejtben azonosak, ami megint nemcsak azt
bizonyítja, hogy ezek nagyon régi eredetű reakciók, hanem azt is, hogy
a ma élő sejtek közös őstől származnak.
A sejtek az evolúció során válságok és azok megoldása alapján
fejlődtek. A legtöbb válságot prokarióták élték át, hiszen ők jelentek
meg először Földön. Ennek következtében a prokarióták mutatják a
legnagyobb változatosságot a biokémiai reakciókban.
Az első válság akkor következett be, amikor a környezetből elfogytak
az építőkövek és az energiaszolgáltató vegyületek (szénhidrátok). A
kismolekulájú anyagokért való versengés felgyorsulásával azok a sejtek
jutottak előnyhöz, amelyek a légkör CO2-ját mint szénforrást
hasznosítani tudták (autotróf szervezetek). A környezetben CO2 nagy
mennyiségben volt jelen, de a hasznosításához energiára volt szükség,
hiszen a CO2 a szénnek a legoxidáltabb formája, és a sejtfelépítéshez
redukálni kell. A sejtek a CO2 hasznosításához az energiát fényből
nyerték (fotoszintézis), egy olyan pigment-molekula (klorofill)
kialakításával, melynek elektronját a fény gerjeszteni képes. A
legelső fény hajtotta reakció a CO2 redukciójához szükséges redukáló
erő előállítása volt a Földön. E reakció során az elektron egy gyenge
elektrondonortól egy erős elektrondonorra vándorol a klorofill által
befogott fényenergia segítségével. Az erős redukálószer pedig
redukálja a CO2 -t. A legelső elektrondonor a H2S lehetett; ekkor a
folyamatban elemi kén keletkezett:
6CO2 + 12 H2S ------> C6H12O6 + 12 S + 6H2O
Ez a folyamat először a zöld-kénbaktériumokban alakult ki.
A baktériumok egy másik csoportja az energiaválság leküzdésére
felfedezte a napenergia hasznosítását és így kialakultak a fototróf
szervezetek. Klorofill tartalmú pigmentek segítségével kidolgoztak egy
elektrontranszport rendszert, amelynek segítségével a fényenergiát
kémiai energiává (ATP szintézis) alakították.
A cianobaktériumok CO2-t és N2-t fixálnak
A cianobaktériumok (vagy másnéven kékes-zöldalgák) egyesítették és
tovább tökéletesítették a kétféle bakteriális fotoszintézis előnyeit
és ennek eredményeként a H2O lett az elektrondonor és a folyamatban O2
keletkezett a fotoszintézisük eredményeként:
6CO2 + 12 H2O -------> C6H12O6 + 6O2 + 6H2O
Az O2 felszabadításával járó fotoszintézis óriási jelentőségű volt,
mivel ehhez a folyamathoz minden szükséges alapanyag bőségesen
rendelkezésre állt, és ezért az élet felvirágzott. Ebből arra
gondolhatnánk, hogy nem is volt több válság!
Ekkor jött azonban a nitrogén válság, mert a szerves nitrogén források
elapadtak. Molekuláris nitrogén rengeteg volt a légkörben, azonban ez
nagyon nehezen lép reakcióba. A cianobaktériumok azonban ezt a
problémát is megoldották és kidolgozták a N2-fixálást folyamatát,
amelyben nitrogenáz enzimet használnak. Ebben a folyamatban rengeteg
energia árán ammóniává redukálják a molekuláris nitrogént. A
cianobaktériumok még ma is a szén és a nitrogén fő fixálói; ezek a
legönfenntartóbb organizmusok a Földön, hiszen vízen és levegőn (CO2
és N2) képesek élni és ezenkívül csak napenergiát igényelnek.
Kialakulások következtében azonban a földi feltételek alapvetően
megváltoztak.
A baktériumok a tápanyag molekulák aerob oxidációjára képesek
Régen nem az ember, hanem más organizmusok csináltak forradalmi
változást a környezetben. Nagyon fontos azonban, hogy ezek a
változások sokkal lassabban következtek be, mint manapság az emberiség
által okozott környezeti változások. A környezeti változások sebességi
aspektusa azért fontos, mert a lassú változásokhoz az élővilág
alkalmazkodni tud, a gyors változások viszont elpusztíthatják az
életet.
Az emberiség megjelenése előtt a legjelentősebb környezeti változás
valószínűleg az oxigén megjelenése volt a cianobaktériumok működése
következtében. Mivel az élet O2 mentes (anaerob) környezetben alakult
ki, és az oxigén nagyon reaktív, miáltal a sejtalkotókkal reakcióba
lép, ezért toxikus hatású volt az első élőlényekre. Ezt bizonyítja a
mai anaerob sejtekre gyakorolt hatása: az anaerobok (melyek nem
rendelkeznek e téren semmiféle védekező mechanizmussal) O2
jelenlétében azonnal elpusztulnak. Az O2 megjelenése tehát újabb
válságot okozott a Földet akkoriban benépesítő anaerob sejtek számára
(O2 válság).
10. ábra: Az O2 koncentrációjának alakulása a Föld atmoszférájában.
Az oxigén megjelenését követően az anaerob sejtek az alábbi
lehetőségek közül választhattak, hogy elkerüljék az O2 toxikus
hatását:
- olyan anaerob környezetbe bújtak, ahol nem volt oxigén,
- kifejlesztettek egy enzimes védekező mechanizmust az O2 károsító
hatásával szemben és kialakultak az aerob baktériumok.
Az O2-nek azonban nemcsak negatív, hanem pozitív hatásai is voltak az
élet földi fejlődésére:
- Az O2-es légkör felső határán kialakult az ózon réteg és kiszűrte a
Nap UV sugarait. Az oxigénmentes légkörben az UV sugárzás elől a vizek
alsóbb rétegeibe bújt sejtek meghódíthatták a vizek felső rétegeit és
a szárazföldet is.
- Az O2 hallatlan reaktivitása révén megteremtette a vegyületekben
lévő kémiai energia sokkal teljesebb kiaknázásának lehetőségét, és nem
csoda, hogy a sejtek ezt a lehetőséget ki is használták. Az O2
lehetővé teszi az energiaszolgáltató molekulák CO2-á és H2O-é történő
sokkal teljesebb elégetését, aminek eredményeként sokkal több energia
szabadul fel. Ezt a folyamatot légzésnek (respirációnak) hívjuk és a
fotoszintézishez hasonló mechanizmussal ATP szintézist eredményez. A
bakteriális fotoszintézisben kialakult elektrontranszport lánc
módosításával kialakult az aerob sejtek légzési elektron transzport
lánca.
Az eukarióta sejtek kialakulása
1.5 milliárd évvel ezelőtt egy újabb mérföldkő következett be a sejtek
a fejlődésében. A prokarióta sejtek egy csoportja ugyanis nemcsak a
biokémiai reakcióit “finomította”, hanem az egyszerű belső struktúrája
vált komplexebbé. A DNS a prokariótákban nagyon gyakran a citoplazma
membrán környezetében helyezkedik el illetve ahhoz kapcsolódik. Egy
ősi prokarióták egy csoportjában valamilyen oknál fogva a
plazmamembrán befűződött, és a DNS-t kettős membránnal ölelte körül. A
citoplazma membrán ily módon történő lefűződésével jöhetett létre a
valódi sejtmag illetve annak kettős membránja és kialakultak az ún.
eukarióta (valódis sejtmagvas) sejtek (eu = valódi).
11. ábra: Az eukarióta sejtek sejtmajának feltételezett kialakulási
mechanizmusa.
Az eukarióták gazdag belső membránstruktúrával rendelkeznek
Az újonan kialalkult eukarióta sejtekben a sejtmagmembrán a DNS
számára védelmet biztosított és így annak mennyisége jelentősen
növekedhetett a prokarióta sejtekhez képest. Egy humán sejt például
1000-szor annyi DNS-t tartalmaz, mint egy baktérium. A DNS
mennyiségének gyarapodása a sejtek térfogatának növekedésével is
együtt járt. Az eukarióta sejtek kb. 100-szor nagyobbak, mint a
prokarióták. A sejtméret ilyen mértékű növekedése azonban számos
problémát okozott az eukarióták számára. A nagyobb sejtméret ugyanis
nagyobb sejtfelületet is igényel, mert:
- az anyagok a plazmamembránon keresztül lépnek be a sejtbe,
- a membrán ezen kívül fontos biokémiai reakcióknak (lipid szintézis,
légzési elektrontranszport stb.) is a helyszíne.
Ha gömbszerűnek képzeljük a sejtet, akkor könnyen belátható, hogy a
méret (a gömb sugara) növekedésével a térfogat köbösen növekszik, míg
a felület csak négyzetesen. Ez azt jelenti, hogy a méret növekedésével
a fajlagos felület csökken. Ezért, hogy a nagy eukarióta sejtek
megtartsák a prokariótákra jellemző nagy fajlagos felület arányt,
belső membrán struktúrákat alakítottak ki.
Ennek első lépése az lehetett, hogy a sejtmaghártya egy labirintus
alakú kompartmentet, az ún. endoplazmás retikulumot (ER) alakított ki.
Az ER membránján lipidek és fehérjék szintézise történik. Az
endoplazmás retikulumból lesarjadzó vezikulumokból képződtek a Golgi
apparátus lapos zsákjai, amelyek az ER-ben szintetizált molekulák
módosítását és továbbítását végzik.
Felvetődik a kérdés, hogy a belső membrán-struktúrák kialakulása
miként oldja meg a fajlagos felület csökkenésének előbb említett
problémáját? Erre a kérdésre az a válasz, hogy ezek a belső membránok
állandó és dinamikus kapcsolatban állnak a citoplazma membránnal.
Nevezetesen, a Golgi zsákjaiból folyamatosan vezikulumok hasadnak el
és olvadnak össze a sejtmembránnal, miközben tartalmukat a sejt
környezetébe ürítik. Ez a folyamat az exocitózis, amelynek révén a
sejt belsejében elhelyezkedő membránnal határolt organellumok a sejt
környezetébe tudnak anyagot exportálni és a sejtfelszín növelését is
szolgálják. Az eukarióták membránnal határolt vezikulumokkal nemcsak
anyagot tudnak exportálni, hanem a környezetükből anyagot is képesek
felvenni. Ez az exocitózissal ellentétes folyamat az endocitózis,
melynek során a citoplazmamembrán egy darabja befűződik, leszakad és
egy citoplazma-membrán határolta vezikulumot hoz létre, ami külső
anyagot vagy a sejtfelszínen megtapadt anyagot tartalmaz. Ezzel a
módszerrel bizonyos eukariótasejtek egészen nagy részecskéket (pl.
idegen sejteket) is képesek bekebelezni (fagocitózis). Természetesen
ezeket a sejt környezetéből felvett anyagokat meg kell emészteni, ami
ugyancsak membrán határolt organellumokban, a lizoszómákban történik,
melyek emésztőenzimeket tartalmaznak. Szintén membrán határolja az
eukarióták peroxiszómáit is, melyeknek a H2O2 termelésben van
szerepük. Ezek a fentebb tárgyalt belső membrán-struktúrák egy
különálló kompartmentet alkotnak az eukarióták citoplazmájában, és a
sejt térfogatának felét is elfoglalhatják. A citoplazma maradék részét
citoszólnak nevezzük.
A mitokondriumok kialakulása
Az oxigén megjelenését az eukarióta sejtek őse úgy vészelte át, hogy
szimbiózist alakított ki egy aerob baktériummal; így alakult ki az
eukarióták mitokondriuma. Ez a Lynn Margulis-tól származó ún.
endoszimbiózis elmélet a legkézenfekvőbb magyarázat az eukarióta
(valódi sejtmagvas) sejtek keletkezésére.
1
2. ábra: A mitokondriumok kialakulása az endoszimbiózis elmélet
szerint.
Ennek megfelelően a mitokondriumok felelősek az eukarióta sejtek
légzéséért. Az endoszimbiózis elképzelést számos bizonyíték támasztja
alá:
- a mitokondriumok mind a méretüket, mind az alakjukat tekintve nagyon
hasonlók a ma élő baktériumokhoz és osztódásra képesek.
- saját DNS-el rendelkeznek, amely szerkezetét tekintve hasonló a
prokariótákéhoz.
- saját fehérje-szintetizáló apparátussal rendelkeznek, mely szintén a
prokarióta apparátushoz hasonló.
A legmeggyőzőbb bizonyítékot a mitokondriumok endoszimbiózisos
eredetére azok a köztes állapotok jelentik az eukarióta fejlődésben,
amelyek még ma is fellelhetők:
- Vannak ugyanis olyan ősi eukariótára hasonlító ma élő
eukariótasejtek, melyek oxigénszegény környezetben (bélben) élnek és
nincs mitokondriumuk (pl. diplomonad).
- Az amőba Pelomyxa palustris-nak pedig nincs mitokondriuma mégis van
oxidativ metabolizmusa, mert a citoplazmájában baktériumok élnek.
Az aerob baktériumokban a plazmamembrán felelős a légzési energia
termelésért. Az eukariótákban a mitokondrium átvette ezt a funkciót a
citoplazmamebrántól, ezért a plazmamembrán új feladatokat láthat el.
A kloroplasztok is bekebelezett prokarióták leszármazottai
A kloroplasztok a cianobaktériumokhoz hasonló O2 felszabadulással járó
fotoszintézist végeznek, ezért nem megdöbbentő, hogy ezek bekebelezett
cianobaktériumok leszármazottai.
Nagyon fontos azonban, hogy a mai eukarióták mitokondriumai és
kloroplasztjai sok mindenben különbözik a ma élő aerob baktériumoktól
és cianobaktériumoktól:
- DNS-ük is kisebb, mint a baktériumoké,
- sok molekulát nem maguk állítanak elő, hanem készen kapják a
gazdasejttől.
Ezen különbségeknek az a magyarázata, hogy mindketten szimbiózis
eredményeként baktériumokból erednek, de nagy evolúciós változáson
mentek keresztül, és ma már nagy részben a gazdasejttől függenek.
Emellett a mitokondriumok általánosak az eukariótákban, kloroplaszt
viszont csak növényi sejtekben fordul elő.
Az eukarióta sejteknek belső váza van
Minél a nagyobb a sejt és minél strukturáltabb a belseje, annál
fontosabb ezeknek a struktúráknak a megfelelő helyen való tartása és
esetleges mozgatása. Minden eukarióta sejtnek van citoszkeletonja, ami
a sejtnek alakot biztosít, a mozgás képességet adja, az organellumok
elhelyezkedését és mozgatását biztosítja.
A citoszkeleton fehérje fonalakból (filamentumokból) áll. A két
legfontosabb típus: aktin filamentumok és a mikrotubulusok. Ezek a
fehérjefonalak nagyon régi képződmények lehetnek, mert szinte
változatlanok az eukariótákban. Mindkét filamentum kulcsszerepet
játszik a sejtek ill. a szervezetek mozgásában; az aktin az izomban, a
mikrotubulusok pedig a csillókban és ostorokban találhatók meg. A
belső mozgásokban is szerepük van: mikrotubulusok alkotják a mitózisos
orsót. Enélkül az eukarióta sejt nem tud szaporodni.
Az eukarióta setekben a genetikai anyag igen komplex módon csomagolt
Az eukarióták igen sok DNS-t tartalmaznak (egy humán sejt 1000-szer
többet, mint egy baktérium). A DNS hossza olyan nagy, hogy nagy a
törés és szakadás veszélye. Ezért jól be van “csomagolva”. Az
eukariótákra jellemző fehérjék a hisztonok, amik a DNS-hez kötnek, és
“becsomagolják” őket kezelhető kromoszómákba. A DNS kromoszómákba
“csomagolása” előfeltétele a sejtosztódásnak. Kevés kivételtől
eltekintve minden eukariótában vannak hisztonok és ezek nagyon
konzervatív fehérjék .
A DNS-t az eukariótákban magmembrán is védi. A genetikai információ
kifejeződésének két fő lépése igy térben elválasztódik:
1. DNS transzkripciója (átirás)
2. RNS transzlációja (átforditás)
A prokariótákban ezek a folyamatok nincsenek térben elválasztva: az
RNS szekvenciák transzlációja azonnal megindul, amint az átiródott
(mielőtt a szintézise befejeződött volna). Az eukariótákban a
transzkripció színtere a sejtmag, míg a fehérjeszintézisre a
citoplazmában kerül sor. Mielőtt az RNS elhagyná a sejtmagot érési
folyamaton (RNA processing) kell keresztülmennie, melynek során egy
része kivágódik, más része pedig módosul.
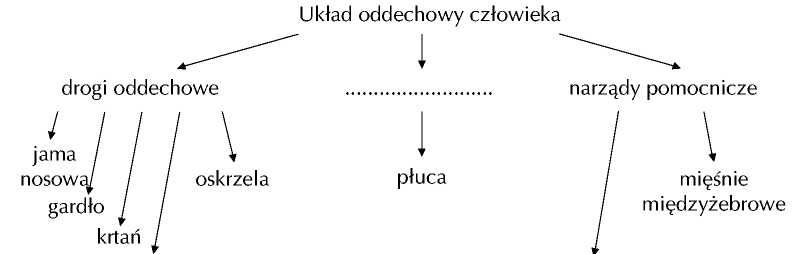 KRWIONOŚNY I ODDECHOWY 1UZUPEŁNIJ SCHEMAT TCHAWICA PRZEPONA 2 ZAZNACZ
KRWIONOŚNY I ODDECHOWY 1UZUPEŁNIJ SCHEMAT TCHAWICA PRZEPONA 2 ZAZNACZ ÖĞRENCILER IÇIN ISTENEN BILGILER (UYGUN FAALIYET GÖRE DOLDURULUR) HAYATBOYU
ÖĞRENCILER IÇIN ISTENEN BILGILER (UYGUN FAALIYET GÖRE DOLDURULUR) HAYATBOYU APPLICATION FOR AREA REPRESENTATIVE STATUS THIS FORM IS TO
APPLICATION FOR AREA REPRESENTATIVE STATUS THIS FORM IS TO AJUNTAMENT DE LA FONT DE LA FIGUERA D ESTEBAN
AJUNTAMENT DE LA FONT DE LA FIGUERA D ESTEBAN DIOCESE OF LEEDS DBI SECTION 48
DIOCESE OF LEEDS DBI SECTION 48  CUERDAS SECANTES Y TANGENTES EN UNA CIRCUNFERENCIA TEOREMA 1
CUERDAS SECANTES Y TANGENTES EN UNA CIRCUNFERENCIA TEOREMA 1 ZADANIE 1 DRUŻYNA MA DO DYSPOZYCJI MOZAIKĘ FIGUR ORAZ
ZADANIE 1 DRUŻYNA MA DO DYSPOZYCJI MOZAIKĘ FIGUR ORAZ APE TURNOS PARA LA PRESENTACIÓN DE SOLICITUDES SE COMUNICA
APE TURNOS PARA LA PRESENTACIÓN DE SOLICITUDES SE COMUNICA O FFICE OF FINANCIAL AID PALATKA CAMPUS 5001 ST
O FFICE OF FINANCIAL AID PALATKA CAMPUS 5001 ST