fisiologia vegetal la célula vegetal estructuras similares a la célula animal. diferencias: cloroplastos, gran vacuola central y pared
FISIOLOGIA VEGETAL
La Célula Vegetal
Estructuras similares a la célula animal.
Diferencias: Cloroplastos, gran vacuola central y pared celular.
Sistemas De Membrana
REL: Da origen a la mayoría de los sistemas de membranas celulares.
RER: Relacionado con la síntesis de proteínas.
Ap. Golgi: Estructura polarizada formadora de vesículas, que se forman
en una cara y se expulsan por la otra. Remodifica moléculas, sintetiza
algunos polisacáridos de la pared. Se cree que es también un sistema
para eliminar substancias que impiden el buen funcionamiento de la
planta.
Vacuola: Estructura de mayor tamaño. Cuanto mayor la célula es mas
madura. Tiene membrana sencilla (tonoplasto). Disolución diluida de
ácidos orgánicos, iones, proteínas en poca cantidad y azucares.
Función: mantiene el volumen celular y la turgencia. Crecimiento
celular. Durante la senescencia muchos enzimas hidrolíticos están en
la vacuola. Homeostasis, mantenimiento de niveles, lo que no usa lo
incluye en la vacuola. El pH de un limón es 3 pero el metabolismo se
da en un pH de 6.5-7.0 por lo que dentro de la vacuola es 3 pero en el
citoplasma es 7. acumula tóxicos y colorantes de las plantas.
Almacenamiento de proteínas.
Pág. 5 Cloroplasto: doble membrana (teoría del endosimbionte), DNA
propio. Tienen unas estructuras membranosas, los tilacoides (bolsitas
de membrana externa con un espacio interior). Los grana son
acumulaciones de tilacoides. Los tilacoides pueden ser de dos tipos:
Tilacoide del grana = lamelas del grana
Tilacoide del estroma = están fuera del grana interconectando los
grana (estroma es la matriz donde están los tilacoides).
Es un orgánulo que permite la absorción de energía luminosa y la
conversión en energía química. Las reacciones no luminosa se dan en el
estroma.
Mitocondria: doble membrana y DNA propio. Tiene una serie de crestas
de membrana interna lo que le permite compartimentalizar lo q se usa
para crear gradientes y usar bombas de protones para generar ATP
gracias a una membrana interna impermeable.
Micro cuerpos: peroxisomas y glioxisomas. De membrana sencilla.
-Peroxisomas: descomponen el H2O2 mediante la catalasa. Por las
reacciones fotosintéticas o por estrés el H2O se transforma en H2O2
(toxico) y la súper óxido dismutasa y la catalasa lo transforman en no
toxico. Fotorespiración y degradación de lípidos.
Plasmodesmos: interconexiones entre células vegetales. La pared esta
interrumpida y se conectan los citoplasmas, gracias al desmotúbulo, q
es una vesícula de RER aplastada q conecta dos células atravesando el
plasmodesmo.
PAG 6 Ø De 60 aunque solo la atraviesan moléculas de 2 NM pues hay
proteínas estabilizando el canal y ponen trabas al paso.
Citoesqueleto: red tridimensional q hace una estructura interna
determinada. Función = transporte de vesículas. Organización espacial
de la célula. Diferenciación. Movimientos del citosol.
- microtúbulos proteína globular (tubulina) de dos tipos, y β. Se
unen para dar un heterodímero. Cuando el nivel de Ca 2+ baja, se unen
los heterodímeros formando un tubo hueco. Es polarizado. Cuando el Ca
2+ aumenta se separan los heterodímeros.
- microfilamentos = dos filamentos de actina q se asocian de forma
helicoidal. Estructuras polarizadas de vida corta.
- filamentos intermedios = láminas nucleares (péptidos en la parte
interna de la membrana nuclear) y queratinas (función del
Citoesqueleto). Estos polímeros son repeticiones de un monómero q
puede unirse de forma paralela o antiparalela.
Organización del Citoesqueleto
5 tipos de organización.
- Organización cortical = en la superficie celular tendencia a
depositar microtúbulos. Tuene q ver con la deposición de celulosa.
- Formación de la banda de la preprofase = antes de la profase se da
Pág. 7) una deposición de microtúbulos q indica a la célula como ha de
dividiese. También indica donde se tiene q depositar la pared para dar
lugar a la zona de separación.
- Huso mitótico = los cromosomas se mueven por unos canales q son
parte del Citoesqueleto.
- Formación del fragmoplasto = son vesículas q vierten material de la
pared para formar una pared intermedia.
- Organización radial = permite la organización de la estructura
interna de la célula.
Plastidios
Orgánulos de las célula, se dividen en:
- leucoplastos = plastidios sin color, amiloplastos, proteinoplastos.
- cromoplastos = plastidios q acumulan pigmentos q dan color a la
planta.
- cloroplastos = doble membrana, tienen su propio DNA, plastidios
verdes en donde se da la fotosíntesis.
Todos provienen de un protoplastidio inicial, y dependiendo de la luz,
función, etc. se transforman en un tipo o en otro. Se pueden
interconvertir unas en otros (leucoplastos de la patata cuando les da
la luz pasan a cloroplastos). Etioplastos son cloroplastos inmaduros.
Pared celular
Necesita mantener intercambio y ser citoesqueleto.
- Celulosa = compuesto mas abundante, glucosas unidas por enlace
β(1-4) insoluble en agua, hidrofílico, resistente al ataque de enzimas
a pH fisiológico. Son enlaces débiles pero muy abundantes lo q da gran
estabilidad y rigidez. Las cadenas de glucosa se van depositando en
forma de entramado.
- Hemicelulosa = 2º más abundante. Cadenas de glucosa con algunas
ramificaciones de otros azucares (xilulosa, galactosa, fucosa).
Estas cadenas no se empaquetan.
- Calosa = hemicelulosa especial q se forma dentro de la planta.
- Pectina = propio de dicotiledóneas. Cadenas de Ac . galacturónico q
en un punto tienen otro azúcar actuando como discontinuidad (ramnosa).
Están estabilizadas por iones Ca2+ . la parte externa de las células
esta cimentada de pectinas.
- Proteínas = Extesina = 10%, se suponía q era responsable de la
elongación celular. Es especial pues no cumple las propiedades de la
proteína. No se digiere bien por proteasa. Pobre en diversidad de aa.
(prolina (45% hidroxiprolina)).
Las proteínas pueden ser estructurales o enzimáticas de dos tipos:
Glucosidasas =para romper la pared. Función de defensa.
Oxidorreductasas = una la peroxidasa. Función defensiva y formación de
ciertos compuestos como la lignina.
- Lignina = formado por fenoles sencillos procedentes de la ruta del
Ac. Shikimico (anillos bencénicos). Precursores de la ruta son
eritrosa 4P y PEP q dan DHAP q dará Trp, Tyr y Phe, a partir de los
cuales salen el resto de aa. La enzima es la PAL (fenilalanina
amonioliasa). Del Phe se saca el Ac. Cumärico, ferúlico y sinápico y
de aquí se dan los alcoholes cumarílico, coniferílico y sinapílico q
por deshidrogenaciones y condensaciones dará la lignina. La lignina
solo aparece en la pared secundaria y se da en células q no son
metabolicamente activas. Función defensiva contra patógenos (sus
monómeros son tóxicos).
- Suberina = ácidos grasos y fenoles.
- Cutina = capa de protección de cera formada por ácidos grasos.
- Metabolitos 2º = fenoles alcaloides y terpenoides.
Localización de los componentes de la pared celular
En los polisacáridos q :no son celulosa los monómeros se dan en el
citoplasma (Ap. De Golgi) y se vierten al exterior.
La deposición de las fibras de celulosa la da el complejo celulosa
sintasa (roseta de 6 unidades con un hueco en medio).
Las rosetas avanzan dejando una serie de glucosas q se ensamblan entre
si tras su síntesis. Según se da la pared se van formando
microfibrillas de celulosa q al unirse a la hemicelulosa y demás
componentes se va estabilizando. También se unen material péctico y
proteínas.
En medio de la célula para la división se va a dar el fragmoplasto q
hace q vesículas del Aparato de Golgi segreguen material de la pared
para dar la placa ecuatorial o placa celular. Los microtúbulos (MT)
cambian de orientación dirigiéndose a los laterales para llevar
vesículas a las células hijas.
Hay dos capas en la pared. Todos tienen pared 1ª pero la 2ª solo
algunas. Pared 1ª = compuesta por la pectina de la lamina media,
proteínas, celulosa no muy ordenada, hemicelulosa. La tienen células
metabolicamente activas. Pared 2ª = compuesta por celulosa muy
engrosada y ordenada, deposiciones de lignina. Se sintetiza después de
la 1ª y tiene función de sostén. En células inactivas.
La Banda de Caspary esta en la endodermis y obliga al flujo de agua a
ir en una dirección establecida.
Funciones y procesos relacionados con la pared celular
- Regula el intercambio celular (iones, gases, minerales y sustancias
orgánicas). No puede estar lignificada pues no pasaría el agua.
- Soporte esquelético q contrarreste la fuerza del agua.
El primer problema a soportar por las células hijas es la rigidez de
la estructura q impide aumentar su tamaño.
Hay 5 tipos de enzimas = auxinas (AIA), citoquininas, giberelinas,
ácido abscísico, etileno (gas), otros).
Auxinas (AIA, IAA) = estimulan la elongación, disminuye el nivel de
hemicelulosa. Las Glucosidasas tienen un pH optimo ácido. Las AIA
tienen dos efectos:
- Rápido = activa la ATPasa q bombea H+ al medio externo lo q baja el
pH lo q activa las Glucosidasas q atacan los polisacáridos no
celulósicos. La pared se hace menos rígida y para contrarrestar los
protones entran cationes y por osmosis agua lo q hincha la célula.
- Lento = activa enzimas q activan genes q codifican proteínas para la
síntesis de polisacáridos de la pared. También se da la síntesis
Las auxinas son fenoles del ciclo del Ac. Shikimico a partir del
triptófano. Implicadas en la extensibilidad de la pared celular. Papel
en el fototrofismo.
Las plantas son mas verdes con baja iluminación, crecen más del lado
no iluminado (para doblarse) las células son mas grandes.
Papel en el gravitropismo para la posición de las raíces.
Abscisión foliar y desarrollo del fruto. La abscisión se produce por
una situación de estrés q produce la rotura de la pared celular en
determinadas células. La célula produce continuamente enzimas de
degradación. Si la relación AIA/etileno es alta la hoja no se cae.
Estas enzimas también tienen q ver con la maduración de frutos (una
fruta madura cuando las paredes celulares se van degradando), una
hormona inductora es el etileno.
Sustancia de reserva. La pared puede servir como almacén en ciertas
condiciones. Conversión de polímeros estructurales en polímeros de
reserva (galactomananos q con agua forman un gel q evita la
desecación, xiloglucanos, galactanos).
Defensa ante estrés abióticos
El estrés lo producen: desecación, salinidad, metales pesados,
temperatura, luz (alta iluminación, UV)
La celulosa aparece en mayor cantidad, hojas coriáceas con cutícula
engrosada. Se incrementan los niveles de extesina y de calosa q se
acumula en los vasos impidiendo q circulen los fluidos. Según aumenta
el estrés : aumentan los depósitos de lignina y suberina en hojas,
cutina en hojas para impermeabilizar y producción de fenoles
sencillos.
Interacción con microorganismos (Rhizobium)
La pared es una barrera física. Por lo q hay diferencias con
microorganismos (MO) simbióticos o patógenos.
- Simbiosis = el 90% de las plantas están micorrizadas (planta
-hongo). Prácticamente ninguna planta vive aislada (Rhizobacterias).
La planta obtiene como beneficio el movimiento de los nutrientes
provocado por la bacteria. La bacteria los compuestos q excreta la
planta.
Proceso de simbiosis: Preinfección: proceso químico, las uniones se
dan en condiciones limitantes por lo q la planta ha de estar
preparada. Reconocimiento: proceso químico. Colonización: la
endodermis no la pueden atravesar los MO. La bacteria genera enzimas q
degradan la pared, la célula se invagina y penetran los MO pero no al
citoplasma. Para q se de la infección la planta emite flavonoides q
activan los mecanismos de Rhizobium (activa genes) q dan una molécula
q da la formación de nódulos es la planta donde se forman los nuevos
Rhizobium. No se rompe la membrana para q el MO entra, sino q se da
una invaginación hasta q queda rodeado. La bacteria tiene capacidad de
tomar N y transformarlo en amonio.
Las micorrizas son unas estructuras en forma de Y con pelillos gracias
al hongo, lo q le permite explorar cada partícula del suelo. Esta
asociación solo se da cuando los dos tienen algún déficit.
- Patógenos = provocan en la planta un estrés biótico. Hay dos
barreras. Mecánica : una pared muy entramada da problemas para la
entrada del MO. Fisiológica : la respuesta a la infección puede ser
localizada o sistemática. Localizada si se da justo donde se da la
infección. Sistemática si la planta al ser infectada adquiere una
información SAR (resistencia sistemática adquirida), el Ac. Salicílico
es el responsable ya q su elevado nivel indica a los tejidos q hay
infección además se puede volatilizar indicando a otras plantas
peligro (se metila para hacerse volátil).
Como se entera la planta de la infección : los genes R codifican los
receptores de membrana a donde se unen los inductores (oligosacarinas
= prot. lip. ) y ponen en funcionamiento el mecanismo de defensa
cuando el patógeno se acerca hay enzimas hidrolíticas q degradan pared
y son estos restos de pared los q informan de un proceso de infección.
Las oligosacarinas provienen de la pared del MO y de la propia célula.
El mecanismo de defensa son una serie de respuestas:
- Hipersensitivas = la planta responde exageradamente a la presión.
Las células donde se ha dado la infección se mueren formando una
costra q protege al resto de los tejidos.
- Respuestas fisiológicas = Fitoalexinas: actividad antimicrobiana,
son fenoles sencillos y algún terpenoide. Medicarpina, gliceolina,
capsidiol.
Explosión oxidativa: se acumulan intermediarios tóxicos como H2O2
radicales O-2 y OH para destruir al patógeno.
Incremento de la barrera física: reforzamiento mecánico de la pared.
Aumento de enzimas hidrolíticas: gluconasas y quitinasas.
Aumento de proteasas:
Aumento de la acción de auxinas: incremento del metabolismo fenólico q
aumenta el AIA q produce nuevos tejidos.
Proteínas RIP: inhibidores de ribosomas . glucosilan el RNAr de los
ribosomas eucariotas de algunos hongos lo q les imposibilita para
producir proteínas y acaban muriendo.
El agua como contribuyente de las plantas
El O2 es electronegativo y el H2O forma un dipolo pues el O2 tira de
los e- del H. Debido a la distribución de cargas el O2 interacciona
con otras moléculas (con otros H por enlace débil).
Propiedades fisiológicas de la molécula de H2O
Absorbe poca radiación visible (más infrarroja). Transparente (si no
habría problemas con los pigmentos). Aumenta de volumen al congelarse,
debido a las interacciones q se dan dejando muchos huecos. Alto calor
de evaporación. Alto calor especifico (para subir 1 ºC la Tª de 1 gr.
de H2O se necesita mucha energía). Cohesión – adhesión: cohesión es la
unión entre las moléculas de H2O, adhesión es la interacción con otras
sustancias. Capilaridad es la interacción entre las dos anteriores.
Buen disolvente.
Funciones del H2O en la planta
- Estructural = da la turgencia a la planta. Semilla 5% en la planta
adulta hasta el 95% es H2O.
- Metabólica = es indicador de la actividad metabólica ya q es
disolvente, fuente de iones, gases y e-, además de reactivo.
- En procesos fisiológicos = transporte xilemático y floemático.
Regulación de la Tª.
Factores q condicionan la disponibilidad de H2O por la planta
Importa la disponibilidad del H2O, no la cantidad. Parte depende de la
planta y parte del suelo.
- Potencialidades hereditarias = influye como es la raíz, tallo,
hojas, dimensión de las hojas, relación entre superficie externa e
interna (raiz y parte aérea), estomas y comportamiento.
- Factores externos = el medio hace q el H2O este disponible o no. Dos
niveles : a nivel de suelo: textura, estructura, profundidad,
composición química, pH, aireación, conductividad y capacidad de
almacenamiento de H2O. A nivel de atmosfera : distribución y cuantía
de la vegetación, precipitación / evaporación, energía radiante,
viento, etc.
Factores afectados por el déficit de H2O
Descenso en la absorción de H2O, perdida de transpiración, cierre de
estomas, no se da expansión celular, disminuye la fotosíntesis y la
reducción biológica de N2 y NO3ˉ , menor transporte xilemático.
Todo esto incide en el crecimiento, de manera q el crecimiento es
menor, abundancia cualitativa y cuantitativa de ciertos compuestos.
Relación raiz / parte aérea (la planta invierte más en la raiz para
poder obtener más agua y minimizar la perdida), menor grado de
suculencia. Xilema transporta H2O, floema transporta sabia.
Potencial hídrico Ψ (es negativo)
Magnitud para expresar el estado del agua permitiendo comparar
sistemas diferentes (aire, suelo). Nos da una idea acerca de la
disponibilidad del H2O. El contenido hídrico y el potencial hídrico no
es lo mismo. Viene expresado en bares de presión. 1 bar = 0.987 atm.
El potencial hídrico tiene q ver con el contenido hídrico. De –2 a –8
en mesófilas o hidrófilas. De –8 a –15 bares en mesófilas en sequía.
De –15 a –30 bares en adaptadas a sequía. A –15 bares se da el punto
de marchitez permanente q es el punto a partir del cual la planta no
se recupera. Agua de mar –25 bares. Hay potenciales de hasta –200
bares en las semillas. El potencial hídrico de cada parte de la planta
es diferente, el global de mide en el xilema. En la raiz es de –2.5 y
en las hojas de –2.7 por lo q sube el H2O y en la atmósfera de –30
bar.
Componentes del potencial hídrico Ψ = Ψsolutos + Ψmatricial + Ψpared
La mayoría del H2O celular en la vacuola con solutos Ψs . En
citoplasma y membrana hay H2O y solutos pero no tanta H2O (ambos son
de signo negativo). Por ultimo la pared ejerce fuerza contraria al
H2O, Ψp y es de signo + pues no restringe la disponibilidad de Ψs. Ψm
se desprecia por ser muy pequeño. En los árboles se añade el potencial
gravitacional Ψ g .
Las plantas se recuperan hídricamente durante la noche con un
potencial hídrico mayor (menos negativo).
Potencial de solutos Ψsolutos
En células mesófilas tiene un valor de entre –10 a –20 bar en células
xerófitas puede llegar hasta –100 bar. Para determinar el Ψs :
- Congelar y descongelar para q se rompa la celula. Luego se mide la
concentración de solutos con un psicrómetro.
- Descenso del punto crioscópico. Sabiendo su valor sabremos su
concentración y por tanto su Ψs.
- Otro método es coger una capa de células de cebolla y meterla en
agua destilada (la celula se hincha por entrada de H2O), luego se
añade ClNa al H2O lo q limita su disponibilidad y el H2O sale, lo q
relaja las paredes, se despega el plasmolema de la pared por lo q Ψp =
0 y como Ψm es despreciable tenemos q Ψ = Ψs (punto de plasmolisis
incipiente)
Potencial de pared o de presión o de turgencia Ψp
En mesófilas valores positivos entre 3 (de noche) y 15 (por el día).
Permite a la celula mantener la estructura interna y externa, relación
con las nastias (movimiento de plantas) y fototropismos y
gravitropismos).
El tropismo implica crecimiento, la nastia no. La nastia además es muy
rápida. Para determinar la Ψp :
- En células grandes se inserta un capilar calibrado en la vacuola, de
forma q la presión de la pared introduce parte del contenido vacuolar
en el capilar cerrado.
- En células pequeñas se usa un sensor de presión con un capilar
sensor lleno de aceite.
Potencial matricial Ψm
Representa la adsorción de H2O a los coloides celulares (pared,
almidón, polisacáridos, proteínas. Para determinar el Ψm :
- Se congela y descongela, de manera q nos quedan los tejidos sin el
jugo vacuolar. Introducimos los tejidos en una prensa y la fuerza q se
hace para sacar el H2O es la Ψm .
Se suele despreciar, pero en plantas desérticas, células q están por
debajo del punto de marchitéz incipiente (deshidratadas),o semillas,
no se puede despreciar.
El Ψs en el xilema contribuye muy poco ya q el gradiente de la
transpiración es mucho mayor.
Que le pasa a la planta en situación de sequía
Mesófilas =Evitan la perdida de H2O cerrando estomas, fotosintetizando
a tasas mas bajas, baja su Ψ para poder captar H2O del suelo. Luego
cambia su metabolismo cuantitativamente. Restringe la disponibilidad
de H2O acumulando solutos en la vacuola (prolina, azucares...).
Xerófitas (adaptadas a la sequía)= Distintas estrategias:
- Evadir el problema = son las plantas anuales (con la sequía mueren).
Son plantas de ciclo muy corto con poca biomasa.
- Derrochadoras de agua = plantas de climas muy áridos con raíces muy
profundas para llegar la zona freática (alfalfa).
- Hacer frente a la sequía = plantas suculentas colectoras de H2O,
sistema radical somero, absorben agua del roció, secretan sal,
metabolismo CAM (cactus), hojas espinosas, reflejan la luz, disminuyen
la relación superficie volumen. Acumulan CO2 a la noche en la vacuola
y por el día lo transforman en azucares. Cutícula engrosada y Ψ muy
baja.
- Ahorradoras de H2O = estomas hundidos, hojas pequeñas y deciduas,
pelillos, aumento de solutos, osmorregulación, células muy compactadas
para evitar la evaporación entre ellas. Muchas xerófitas se incluyen
en este grupo y en otros.
- Soportan la sequía = las xerófitas verdaderas cuando hay sequía se
mueren y renacen cuando las condiciones se vuelven optimas. Tolerancia
a la deshidratación, soportan gran perdida de H2O.
Procesos afectados por la sequía
En mesófilas el Ψ comienza a bajar. Cuando una planta esta bien
hidratada la acumulación de Ac. abscísico (ABA) y de solutos es baja.
A –5 bar empiezan a verse afectados ciertos procesos muy sensibles y
se acumula ABA y solutos. Procesos de gran actividad en hidratadas
disminuyen su actividad: síntesis de proteinas, de pares, expansión
celular. A –10 bar se ve afectada la fotosíntesis y la apertura
estomática.
Determinación del estado hídrico de la planta
Para medir el estado hídrico de la planta se mide el contenido hídrico
(Ps es el peso seco, mientras q Pf es el peso fresco).
Otra forma es medir el contenido hídrico relativo CHR (Pt es el peso
turgente. Se toma el tejido y se introduce en agua pata q se hinche en
una atmósfera saturada. Una forma rápida de ver el valor del estado
hídrico es medir la relación
Métodos de medida del potencial hídrico Ψ
Son métodos gravimetritos, se necesita una balanza o una regla para
determinar el Ψ. Se usa en la patata pues es cómodo un corer o
despepitador y se sacan cilindros cortándolos a una longitud
determinada. Calculamos los Ψ se diferentes soluciones de manitol y
sumergimos cada cilindro en una solución diferente, pudiéndose dar 3
casos:
- Si la solución es mas diluida q el tejido por lo q el tejido absorbe
H2O.
- Si la solución tiene un Ψ igual al del tejido no se da intercambio
de H2O.
- Si la solución esta mas concentrada q el tejido saldrá H2O del
tejido.
Método de Chardakov
Se hacen dos baterías de tubos gemelos (mismo Ψ). A cada tubo de una
batería se tiñe con azul de metileno. En los no teñidos introducimos
una hoja y dejamos equilibrar pudiéndose dar 3 casos.
- Q el agua no se mueva,
- Q entre H2O en la hoja.
- Q salga H2O de la hoja.
Luego sacamos las hojas q habrán variado la disolución (o no), tomando
una gota del tubo gemelo teñido y añadiéndola al de la hoja:
- Si la gota se dispersa tendremos la Ψ.
- Si sube la gota es porque es menos densa ósea q la dilución esta más
concentrada porque la hoja ha tomado H2O.
- Si baja es el caso contrario.
Método de la cámara de presión
En una cámara de la q sobresale el tallo seccionado. Se da presión
para llevar la sabia del xilema al punto de corte. Si se necesita
mucha presión, Ψ es muy negativo (seco ). Es un método muy impreciso q
solo vale para la planta.
Métodos de equilibrio de vapor
Se usa el psicrómetro de vapor o psicrómetro isopiéstico. En una
cámara dentro de otra termo estabilizada, con dos cables de distinto
metal, soldados a un termopar. Con la diferencia de Tª se da
diferencia de (energía) en el cable, midiéndose si el termopar se
calienta o enfría. Con el P. De Boyer, el termopar esta en la cámara,
si añadimos una gota de Ψ conocida al termopar, se pueden dar tres
casos:
- El Ψ de la gota es el del tejido: el voltímetro marcara 0.
- El Ψ del tejido es menor el de la gota por lo q el H2O va de la gota
al tejido lo q la enfría y da lectura en el voltímetro.
- El Ψ del tejido es mayor por lo q el agua va a la gota lo q la
calienta y da lectura en el voltímetro.
Los problemas del método son los cambios ambientales, la lentitud
(horas), aunque es muy exacto.
El agua en el suelo
Medio complejo (fase sólida, liquida y gaseosa). El estado hídrico Ψ
del suelo es la suma algebraica de:
Ψ suelo = Ψ sales + Ψ gravedad + Ψ matricial
Ψ sales = La restricción de los iones gases, es muy baja.
Ψ matricial = Debido a sustancias q no se disuelven pero retienen H2O
(restos vegetales, arcillas, etc. Esta Ψ es el componente principal.
Ψ gravedad = Cuando regamos el H2O se pierde por la gravedad. Este es
el H2O gravitacional. La máxima cantidad de H2O q puede retener un
suelo es la capacidad de campo. El H2O se va evaporando hasta el punto
q el suelo ya no puede ceder H2O a la planta, esto es el punto de
marchitez permanente. Por encima de la capacidad de campo la planta no
puede crecer pues nos cargamos la fase gaseosa y la raíz no puede
respirar. El H2O entre la capacidad de campo y el punto de marchitez
permanente es el agua capilar.
Factores q afectan a la disponibilidad de H2O
-Tamaño de las partículas: se clasifican en: Arena gruesa 200 μm –
2mm, Arena fina 20 μm – 200 μm, Limoso 2 μm – 20 μm, Arcilloso menos
de 2 μm. Las partículas tienen espacios gaseosos grandes y el H2O se
pierde. En las partículas finas la fase gaseosa es casi nula.
En un suelo arcilloso la capacidad de campo es de 50% de H2O, en uno
arenoso solo del 15%, (Hoja 38 Fig. 3.23)y los puntos de marchitez son
20% y5% respectivamente lo q muestra q el arcilloso puede perder un
30% de H2O mientras q el arenoso solo un 10% (hay q regar
continuamente). La cantidad de H2O n es importante (para uno
encharcamiento y para el otro sequía). El contenido en sales disminuye
la disponibilidad de H2O.
Conductividad del agua en el suelo
Nos indica el grado de compactación de las partículas del suelo.
Cuanto menos compactado mas se mueven el H2O y los nutrientes.
Rizosfera: es el área del suelo controlada por la raíz.
Suelos artificiales o suelos inertes
Recambiar el suelo es costoso, los monocultivos son antinaturales. El
monocultivo en invernadero lo separamos de sus invasores. Se le pone
un suelo inerte pues los nutrientes se los damos en el H2O. Se le dan
unas condiciones de luz, nutrientes, etc. constantes menos una q se va
variando. (la luz es en cámaras de crecimiento, no se trata de luz
solar).
Tipos de suelos
Les sirven de sostén y para mantener las características hídricas.
- Vermiculita = es una mica calentada a 3.000ºC lo q separa las capas.
Tiene gran intercambio catiónico y puede actuar como reservorio de
nutrientes. Simula suelo arcilloso.
- Turba = restos vegetales tan lixiviados q casi no tienen nutrientes.
Gran capacidad de retención de H2O, simulan suelo poca aireado y
encharcado.
- Perlita = silicato calentado para q se expanda. Cada bola puede
retener 3 o 4 veces su peso en H2O. Bien aireado.
Estos sustratos se combinan entre ellos para dar el tipo de suelo
deseado.
Medición del potencial hídrico del suelo (hoja 39)
- Tensiómetro = columna terminada en una porcelana porosa con H2O. Al
introducirla en la tierra, esta chupara del H2O de la porcelana si
necesita H2O, creando una tensión negativa. Si el suelo esta bien
hidratado marcará 0 o casi nulo.
- Bloque de resistencia eléctrica = dos cables dentro se un bloque de
escayola. Cuanto mas H2O haya en el suelo mayor conductividad y por
tanto menor resistencia.
- Sonda de neutrones = un emisor de neutrones rápidos q los lanza en
todas direcciones, y un contados de neutrones lentos. Como los
neutrones son frenados por los protones cuanto más H2O más se verán
frenados los protones. Sólo vale para zonas con poca MO pues esta
también frena los neutrones.
Absorción y transporte de agua
La absorción y transporte se deben al xilema y floema. El xilema es de
la parte basal a la apical, el floema es de redistribución. El xilema
es tejido muerto y el floema vivo. Todo el tejido muerto en el
interior (xilema) y el vivo en el exterior (floema).
El H2O sube por diferencia de potencial entre la atmósfera y la planta
(hoja 40 Fig. 4.5).
Las paredes están lignificadas y engrosadas.
- Elementos del Xilema = son dos con anatomía y distribución
diferente.
Traqueidas : alargadas y estrechas, presentes en plantas menos
evolucionadas, presentes en gimnospermas. Tienen gran cantidad de
punteaduras.
Vasos :más cortos y bastante mas anchos. Se comunican por las placas
de perforación, llamadas así porque los Plasmodesmos se han abierto
dejando fluir el H2O. Son mas evolucionadas y están presentes en
angiospermas (también tienen traqueidas).
Las células parenquimáticas tienen función de reforzar el sistema
junto a las fibras.
La mayor parte de la madera se debe a restos del xilema. El cambium es
un tejido q diferencia al xilema en el interior, y al floema en el
exterior.
Acompañando al floema siempre habrá células acompañantes con núcleo
(hoja 40).
Las punteaduras surgen de lo q eran los Plasmodesmos. Una punteadura
se separa por la pared celular 1ª q es difusa y porosa (hoja 41) la
pared 2ª por su parte, no deja el paso de H2O, ya q se encuentra
engrosada y lignificada.
Mecanismos del xilema
Básicamente vamos a hablar de dos:
- Presión de raíz : es bombear desde abajo. Indica q hay una fuerza q
tira del agua desde abajo. Gutación= la raíz bombea H2O y hace q el
H2O liquida salga al exterior de las hojas. Se hace a través de los
hidátodos. Se define como liberación de H2O por las hojas debido a la
presión de raíz.
La presión de raíz se da en plantas de pequeño porte, muy bien
hidratadas y q no existe transpiración o q esta sea muy pequeña. Se da
en gramíneas en las primeras horas de la mañana. El H2O tiene q hacer
dos movimientos, uno transversal hacia el xilema y otro a lo largo del
xilema.
Hay células q bombean iones en el xilema por lo q baja el Ψs y entra
H2O lo hace q se bombee por el xilema.
- Teoría de la tensión- cohesión o teoría de Dixon : (Hoja 42 Fig.
4.13) se basa en la diferencia de Ψ entre la atmósfera y la planta por
tanto el H2O pasa de la planta a la atmósfera ya q siempre se evapora
agua.
Esto genera una tensión en el tubo. La cohesión hace referencia a la
capacidad de estar unidas las moléculas de agua por puentes de
hidrógeno.
Al tirar de las moléculas de arriba se tira de las de abajo lo q
provoca la entrada de nuevas moléculas de agua. El H2O puede viajar
por el mismo tubo o por elementos contiguos siendo un movimiento
lateral. Esto crea una gran presión de vacío q puede llegar a colapsar
en época de sequía.
Los tubos tienen paredes semirrígidas de tal modo q no se colapsen.
Antes de q se colapsen, hay un proceso de cavitación o embolismo,
situación donde la fase gaseosa del H2O disuelta en el liquido de la
disolución debido a la presión, pasa a un estado gaseoso formando
burbujas.
Cuando se da una cavitación (hoja 41 fig4.6) la ventaja de tener
placas de perforación es q la burbuja no puede traspasar la placa de
perforación ya q se queda adherida y es tanta la presión superficial q
no puede pasar.
La planta reconoce el problema en el elemento criboso y luego trata de
solucionarlo.
El H2O pasa al elemento criboso contiguo y se evapora.
Por la noche la planta se recupera hídricamente ya q la fuerza de la
transpiración disminuye. El H2O sube y se permite reconstruir las
columnas de H2O. Cuando una planta cavita se puede oír un pequeño clic
lo q nos da idea del numero de cavitaciones de la planta.
Hay procesos q producen cavitación como la congelación y la acción de
algunos patógenos. Los olmos sufren la grafiosis, causada por el hongo
Cerabocisbys olmi. (hoja 43)Este hongo invade los tejidos del xilema y
al crecer provoca q la tensión no sea tan alta y la burbuja pueda
avanzar cavitando todos los elementos criboso. El organismo q
transporta el hongo es el Scolytus multistriatus, cuando este insecto
sale del árbol enfermo lleva esporas y llega a un olmo sano. La hembra
cava un túnel debajo de la corteza y va dejando huevos, las crías
cavaran nuevos túneles alimentándose de la madera, y cuando se
desarrollen extenderán la infección.
La cavitación se extiendo por el xilema y el agua se evapora, las
hojas se secan debido a la oca presión de raíz solo creciendo las
cercanas al tallo.
En un árbol, hay diferente Ψ según midamos a una altura u otra. Cuanto
mas cerca de la base mayor Ψ y cuanto mas lejos menor (mas negativo)
Ψ. A mediodía la Tª es máxima y el Ψ de la atmósfera es mas negativo y
tira del agua de la planta, el Ψ de la planta también disminuye porque
la evaporación es mayor. A mediodía la planta cierra los estomas.
Transpiración y fisiología estomática
Transpiración
Es la perdida del H2O absorbida q tiene la planta en forma de vapor.
El 99% del H2O q absorbe la planta vuelve a perderla. (hoja 44 Fig.
4.9) si miramos un trozo de hoja del exterior al interior tiene
cutícula, por debajo el parénquima en empalizada y a continuación
están las células del mesófilo. En alguna cara de la epidermis hay
discontinuidades q son los estomas q son poros controlados por la
acción de un grupo de células. Se puede evaporar por los estomas o
salir por la cutícula (10% del agua perdida), cuanto mas cerrados
estén los estomas mayor proporción de H2O se pierde por la cutícula.
La capa limite es una especie de aislante q impide la perdida de agua.
Esta capa de aire limite tiene diferencias con el aire externo. La
planta puede desarrollar pelos q aumentan la capacidad de impedir la
perdida de agua.
Según disminuye la humedad relativa el Ψ disminuya. Las plantas
xerófitas tienen modificaciones:
- Desarrollo de cutículas grandes, engrosadas así aumenta la capa
limite.
- Hundir los estomas.
- Desarrollo de pelos para q la capa limite sea mauro.
- Obligar a los estoma q se cierren por el día y se abran por la noche
para evitar el gradiente.
- Células totalmente compactadas sin espacios aéreos, de modo q si el
agua tiene q pasar le cueste mas.
- Disminuir el numero de estomas y el tamaño del poro.
Todas estas estrategias van encaminadas para disminuir el gradiente.
Al aumentar la Tª el Ψ disminuye. El aire frió se satura con poco H2O.
El caliente necesita grandes cantidades de agua para saturarse.
Estomas
Están en todas las partes de la planta excepto en la raíz. La mayor
parte de los frutos son verdes porque fotosintetizan para obtener
carbohidratos, por ello necesitan los estomas para intercambiar gases.
El nº varia de unos 15 – 500/mm2 aunque la mayor parte tienen entre
100 y 300.
Cada especie ha evolucionado de una forma diferente:
- Plantas anfiestomáticas: estomas en las dos caras de la hoja.
- Plantas hipoestomáticas: con estomas en una cara, normalmente en la
parte inferior, excepto las acuáticas q los tienen en la parte
superior.
Estructura de los estomas
Dos formas típicas de estomas (hoja 46 Fig. 4.12). siempre van a estar
rodeadas de células epidérmicas básicamente vacías.
- Estructura de plantas mas evolucionadas propio de gramíneas, con
forma de hueco.
- Estructura de plantas menos evolucionadas con forma arriñonada.
Un estoma consta de dos células q al cambiar de volumen dejan un poro.
Son las células guarda o células oclusivas. En las mas evolucionadas
acompañando a estas células están las células subsidiarias
acompañantes adjuntas o accesorias, q también forman parte del estoma
ayudándole a funcionar mejor.
La mayor parte del intercambio y la evaporación se da por los poros.
El poro se denomina ostiolo y cuando las células están unidas se
encuentra cerrado. Las células oclusivas tienen las paredes engrosadas
en zona concretas. La abertura o cierre del poro se basa en unos
principios q son cuando la célula se hincha se abre el estoma, la
célula se hincha debido a una entrada de agua , el cierre por tanto es
debido a perdida de agua. El citosol esta situado en el extremo de la
célula oclusiva y el medio es pares de manera q cuando se hincha,
debido a la disposición de las fibras deja un hueco.
Las células oclusivas también tienen cloroplastos, mitocondrias,
gránulos de almidón, etc, aunque no tienen plasmodesmos para
comunicarse con las células epidérmicas.
En sus vacuola acumulan grandes cantidades de solutos, sufren cambios
bruscos de volumen debido a la entrada de agua.
Funciones de la transpiración
- Intercambio gaseoso: es un intercambio neto de O2 hacia el exterior,
vapor de agua al exterior y CO2 hacia el interior. Sirve para regular
el intercambio gaseoso con la mínima perdida de agua.
- Regulación de la Tª: pierden gran cantidad de calor con la perdida
de agua, evitan el sobrecalentamiento.
- Concentración iónica: si el agua no se evapora no hay transporte
xilemático y no se puede tirar de los iones.
Mecanismo de funcionamiento (hoja 44 Fig. 3.16)
Se da una acumulación de solutos y esto disminuye el Ψs provocando la
entrada de agua e incrementando el volumen celular, lo q abre el
estoma.
Diferencias entre la célula abierta y cerrada
- Cuando un estoma se abre su Ψs es muy bajo.
- Aumento de la [K+] en la vacuola de las células hasta 8 veces.
- Aumento de malato y sacarosa.
- Gran concentración de Cl-.
- Gran actividad ATPasa y gran salida de protones al medio externo.
Cuando el nivel de CO2 es bajo el estoma se abre.
- Cuando el estoma esta iluminado esta abierto. Se abren al amanecer.
Las plantas crasulaceas hacen lo contrario. Tienen muy difícil la
abertura de los estomas durante el día y los abren al anochecer.
- Cuando se abre el estoma se da un descenso de almidón.
- Si no hay nada de CO2 en el estoma no se puede abrir y si la
concentración en muy alta también se cierran.
Mecanismos de funcionamiento
El pH varia . cuando se esta abriendo se activa una ATPasa q bombea H+
al exterior. Esta basificando el medio, baja el pH en el exterior.
Esto provoca q el almidón se transforme en glucosa q por la glucólisis
da PEP, esto se carboxila a OA y luego se desnitrogena a malato.
El malato es una fuente de H+ para q la ATPasa siga funcionando.
Al bombear H+ al exterior estamos favoreciendo la entrada de ClK. Los
H+ tienden a volver a entrar y arrastran el Cl- al interior q se
estabiliza con el K+ q ha entrado.
El ABA proveniente de las puntas de las raíces o de las hojas, hace q
los estomas se cierren por inhibición de una bomba de protones
dependiente de ATP en la membrana plasmática de las células oclusivas.
Esta bomba normalmente transfiere protones hacia fuera de las células
oclusivas causando un flujo de entrada rápido y acumulación de iones
K+, y luego absorción osmótica de agua y apertura estomática. Sin
embargo, al actuar en el espacio libre de la superficie exterior de
las membranas plasmáticas de las células oclusivas, el ABA cierra la
entrada de K+, por lo q se filtran los K+ y el agua hacia fuera, se
reduce la turgencia y por tanto se cierran los estomas.
A partir de glucosa se sintetiza PEP y rubisco, formando CO2 q vuelve
a malato.
Papel de la sacarosa
Se pensaba q el nivel de K+ subía rápidamente cuando se abría el
estoma, pero no se había planteado si este nivel se mantenía.
Se vio q aunque el estoma seguía abierto la [K+] no se mantenía. A las
11 de la mañana la [K+] desciende y no coincide con el máximo de
apertura estomática.
Al hacerlo con sacarosa al principio no se ve muy estimulada. Según se
va dando la fotosíntesis la concentración de sacaros aumenta siendo
máximo con el máximo de apertura estomática.
La alcalinización del medio es la q desencadena la degradación del
almidón.
Cuando se cierra el estoma la ATPasa se detiene, entran H+
deteniéndose la degradación del almidón y provocando la salida de K+ y
Cl- , el estoma se deshincha, sale el agua, debido a la salida de los
iones.
Factores q inciden en la abertura o cierre de los estomas
- CO2 = la planta iba a perder O2 y agua. La función es captar CO2 q
va al cloroplasto y mediante la fotosíntesis obtenemos carbohidratos.
Cuando el estoma esta abierto entra CO2 creando un gradiente de
concentración. Suponemos q la [CO2] en la atmósfera es 300ppm y la
cámara subestomática 250ppm. La [CO2] en el interior va disminuyendo
porque este CO2 se transforma en carbohidratos. Si se para la
fotosíntesis la concentración interna y externa se igualan y no entra
CO2 .
Si el proceso fotosintético es muy rápido el estoma tiende a abrirse
para mantener el nivel de la fotosíntesis. Si es muy lento el estoma
ajusta su abertura para q no se acumule el CO2 . Es el propio CO2 el q
cierra el estoma si la fotosíntesis no funciona. Si la [CO2] en el
exterior es muy elevada el estoma se cierra para dejar pasar solo el q
necesita.
¿por qué al aumentar la [CO2] el estoma se cierra?
Si aumenta la [CO2] se da un aumento en la aparición de malato, lo q
se traduce en q aumenta rápidamente los protones, bajando el pH
citosólico. Esto inhibe los enzimas de la degradación del almidón
hasta q no se gasta todo el CO2 y vuelve a funcionar todo el sistema.
- Luz : es menos importante q el factor CO2 . una planta iluminada, si
la soplamos cierra los estomas. Se hizo un experimento para ver si el
efecto fotosintético abría los estomas. (hoja 49 Fig. 3.17) Se ilumina
una planta con luz azul, verde y roja. Con luz verde se muere pus no
se absorbe la luz verde. En función de la luz varia la tasa de
fotosíntesis. La luz roja es mas efectiva para realizar la
fotosíntesis. La azul es mas efectiva en cuanto a la conductancia
estomática. Al amanecer las plantas tienden a abrir los estomas sin
fotosíntesis. Es un efecto directo de la luz azul.
La zeaxantina es un pigmento q tiene un papel importante en la
fotoprotección y abertura estomática. Por la mañana hay poca
zeaxantina, a mediodía es máxima y al atardecer disminuye. Midiendo el
contenido de zeaxantina en un célula del mesófilo u en una célula
guarda se ve q en la ultima la concentración de zeaxantina es 4 veces
mayor q en las del mesófilo. Por ello se piensa q la zeaxantina es uno
de los cromóforos responsables de la absorción azul.
Síntesis de la zeaxantina (hoja 50 Fig. 18.26) la zeaxantina se
encuentra en los tilacoides. Para q se forme necesita luz. Se forma a
parir de otra xantofila q es la violaxantina. Esta se acumula de noche
y cuando hay luz se transforma. Cuando hay acumulación de zeaxantina y
se ilumina con luz azul ,se activa, se transforma en otro compuesto
unido a otra proteína y provoca la respuesta de un 2º mensajero q
tiene q ver con el secuestro de Ca2+ al cloroplasto. Esta bajada de
Ca2+ en el citosol activa ala ATPasa. Esto se da en todas las plantas
solo q en algunas esta amortiguado.
En las crasulaceas no se abren los estomas en todo el día. Por la
noche se abren los estomas, entra CO2 y se va fijando en PEP dando OA
q pasa a malato. El malato entra en la vacuola y se acumula.
Durante el día el malato se descarboxila apareciendo CO2 libre q se
combina con la RUBISCO mediante el ciclo de Calvin, fijándolo.
Durante el día las plantas son menos ácidas y por la noche mas ácidas.
Las plantas C3 a la vez q fijan el CO2 lo tienen q gastar perdiendo
agua, por ello son mas sensibles a la sequía.
- Potencial hídrico : un estrés hídrico implica el cierre de los
estomas y es el más importante frente a los demás. Hay 4 sensores
(hoja 49 Fig. 3.29).
Transpiración : sube de día y baja de noche en función del % del valor
máximo. La transpiración y la absorción son diferentes. Durante el día
la absorción es menor q la transpiración y durante la noche es al
revés por ello se recupera la planta.
Hay varios mecanismos por los q el Ψ influye en los estomas:
Mecanismo hidropasivo : para q se abra el estoma la célula necesita
agua.
Mecanismo hidroactivo : el ABA es al responsable del cierre
estomático, es la hormona del estrés y la q sube mas de nivel en una
planta estresada. No importa tanto su concentración como la
distribución. El ABA provoca el cierre estomático evita la perdida de
agua, favorece da conductividad hidráulica del agua en la planta y
capacita para desarrollar nuevas raíces.
El ABA es un terpenoide = isoprenoide, ruta de biosíntesis de
terpenoides, todos son múltiplos de 5, a unidad repetitiva es el
isopentenil pirofosfato (IPP) y la ruta la del acetato – mevalonato.
El IPP proviene del acetil CoA, 3 unidades de este producen un
compuesto de 6C (Ac. Mevalonico) q por descarboxilación forma el IPP.
La síntesis se produce por dos vías, síntesis de novo, o a partir de
un tetraterpeno.
- Oxígeno O2 : muy poco importante ya q su concentración en la
atmósfera es muy grande por lo q habría q variar mucho de
concentración para q afecte. La planta produce O2 pero es un mecanismo
indirecto pues mucho oxigeno cierra el estoma. Si aumenta la
concentración de oxigeno estamos estimulando el proceso
fotorrespiratorio aumentando los niveles de CO2 q provoca un cierre de
los estomas. Las plantas C4 no tienen Fotorespiración.
- Temperatura : influye, aunque no es de los sustanciales. Al aumentar
la Tª se consume CO2 y tiende a abrir los estomas. Hasta 30ºC la
fotosíntesis va pareja con la respiración. A partir de 30ºC la
respiración es superior a la fotosíntesis. Al aumentar la Tª baja la
humedad, evapora mas a la planta llegando a provocar la sequía.
- Velocidad del viento : influye en la transpiración. Cuanto mas
viento reseca mas la planta con lo q al aumentar la velocidad aumenta
la transpiración, obligando a la planta a cerrar los estomas. Influye
indirectamente en la abertura estomática. (hoja 52 Fig. 3.8)Resumen de
todos los factores.
El estrés hídrico domina sobre el resto de los factores. Con alta
iluminación abre mas los estomas q si la iluminación es baja. Las
plantas suculentas durante el día descarboxilan mas CO2 cerrando los
estomas. Una planta C3 con la primera iluminación abre los estomas y
al atardecer los va cerrando hasta q los cierra totalmente por la
noche. Una planta normal a mediodía a veces cierra un poco los estomas
para recuperarse de algún estrés.
Una planta seca intenta abrir sus estomas, pero a eso de las 10 de la
mañana detecta el estrés hídrico. Se activa el mecanismo pasivo
teniendo cerrados los estomas a mediodía cuando la conductancia
estomática tendría q ser máxima.
Medición de la abertura estomática
Se pueden medir dos cosas:
• Si están abiertos o cerrados
• Transpiración
Para ver como están los estomas se visualizan. Para ello se pinta la
superficie vegetal con laca de uñas, con lo q se crea un negativo. Lo
separamos y se mide el poro.
También se pueden usar los porómetros q no indican como esta el poro.
(hoja 53 Fig. 5.9)
- Porómetro de flujo: para plantas anfiestomáticas. Es una especie de
pinza perforada en la q se mete la hoja. Lo q se hace es introducir
aire ejerciendo una presión para ver como pasa el aire de la parte
inferior a la superior.
- Porómetro de difusión: pinza q acaba haciendo una cámara con un
sensor de infrarrojo. Lo q mira es si evapora agua. Cuanto mas abierto
mas difusión de agua. Lo q mide es la humedad del flujo de agua.
Tiene varios problemas: El CO2 absorbe por lo q influye. Para eliminar
el CO2 , el aire se pasa por un filtro q fija el CO2 , de este modo el
aire q pasa es aire limpio.
- Porómetro de inmersión: según la cantidad de transpiración hay una
burbuja q se mueve. La mayor parte de cambios de peso de la planta se
deben a la cantidad de agua. De este modo se puede medir la cantidad
de agua q transpira al día. Esta cantidad de agua es la suma de la
planta y el suelo. Esto es tasas de evapotranspiración. Si solo
medimos la planta es evaporación.
- Lisimetro: permite medir las tasas de evapotranspiración de un suelo
en condiciones medioambientales. Por presión de las bolsas en el suelo
el liquido sube mas o menos según la transpiración. Cuanto mas
transpira menos pesa el suelo
Absorción iónica celular
La absorción se realiza por todas las partes del vegetal. Aunque la
estructura q permite q se de la absorción selectiva es la membrana.
Funciones de las membranas biológicas
• Aislar el medio externo del interno
• Compartimentación del metabolismo
• Regulación de la composición interna
• En la comunicación celular
(hoja 54 Fig. 1) En la membrana hay lípidos (muchos insaturados),
glúcidos y proteínas inmersas en la membrana hay proteínas
distribuidas asimétricamente.
(Fig. 2.4) Las membranas se forman continuamente a partir del RER.
Estas membranas van al Ap. Golgi y de aquí salen vesículas hacia la
vacuola donde se unen y pasan a formar la membrana de la vacuola.
• Gran impermeabilidad selectiva
Muchas moléculas q pasan la pared no pueden atravesar la membrana. Las
acuaporinas son las q dejan pasar al agua.
Hay tres tipos de transporte:
- Pasivo o difusión simple : las moléculas se mueven a favor de
gradiente, de mas concentrado a menos concentrado. Hay dos componentes
q facilitan este movimiento, el potencial químico y el potencial
eléctrico. Si añadimos sacarosa en un vaso de agua, esta se va al
fondo. Al cabo del tiempo la sacarosa se distribuye uniformemente
rompiéndose es gradiente químico generado. Si añadimos ClNa hay q
tener en cuenta el potencial químico y el eléctrico, por eso se habla
de potencial electroquímico.
No utiliza transportadores.
- Difusión facilitada : requieren transportadores. Hay tres tipos de
proteínas q se encargan de este transporte. Carriers, canales y bombas
especificas para el transporte activo. Los tres han de estar en la
membrana para q los iones puedan pasar. El canal es una proteína q
tiene un poro por el q pueden pasar diferentes sustancias q difunden a
favor de gradiente de concentración. Suelen ser exclusivas para un
tipo de ion. Solo esta abierto un periodo de tiempo determinado
abriéndose y cerrando por niveles de Ca2+ hay casos de un canal de
entrada y otro de salida. Los Carriers son un transporte constitutivo
pero mas lento, no gasta ATP, a veces actúa favor de gradiente y otras
veces en contra. La bomba especifica gasta ATP generando un gradiente
de H+ moviendo un carrier.
Características del transporte facilitado.
Siempre se da unido a un transportador de canal o un carrier. No se da
gasto energético. A favor de gradiente. El flujo de transporte siempre
es mayor q la difusión simple. Selectividad de iones. Se da un
fenómeno de competencia de iones (si un canal se parece a otro se dará
competencia entre iones diferentes). Se dan cinéticas de saturación
(sobre todo en los carrier). En alguna ocasión puede darse una
difusión simple en contra de gradiente, es el Potencial de Donan,
(hoja 55) a pesar de ser difusión simple se da una absorción en contra
de gradiente sin gasto de ATP. Para q se de el sistema se necesitan 2
compartimentos separados por una membrana q permita el paso de iones
móviles e impermeable a moléculas cargadas.
- Transporte activo : absorción contra gradiente gastando ATP. Para
ello están las ATPasas q bombean H+ gastando ATP. Se pueden dar dos
tipos de transporte activo:
Primario: se trata de bombear H+ contra gradiente (hoja 56).
Secundario: se utiliza el gradiente de H+ generado en el transporte
primario para movilizar otras moléculas.
• Simporte: las moléculas viajan en el mismo sentido q los H+ . son
moléculas electroneutras.
• Antiporte: las moléculas viajan en sentido contrario. Lo q sale
tiene la misma carga q los H+.
• Uniporte. Intercambio de iones por una despolarización de la
membrana.
Transportadores ABC (ATP binding cassette). Recientemente
descubiertas, estas bombas asociadas al glutatión se localizan en la
vacuola. Precisan hidrólisis de ATP para funcionar. Bombean moléculas
orgánicas. No tienen relación con un gradiente de H+ , las moléculas
se transportan unidas al glutatión. Función: desintoxicación de
moléculas orgánicas toxicas (herbicidas, metales pesados ...).
acumulación de determinados pigmentos en la vacuola. Acumulación de
compuestos antimicrobianos como las fitoalexinas. Evitacion del estrés
oxidativo.
Un ejemplo de estos transportadores son las antocianinas q son
bombeadas por las bombas ABC dentro de la vacuola. Otro transportador
son las fitoquelatinas q se unen a los metales pesados. Las
fitoquelatinas son unas proteínas q se incluyen en el grupo genérico
metalotioneinas. Se distinguen 3 grupos: - Polipéptidos relacionados
con la metalotioneina renal equina. – Polipéptidos no relacionados con
la metalitioneina renal equina. – Fitoqelatinas. Las dos primeras
están codificadas genéticamente, la tercera no. Analizando la
estructura química se ve q es la repetición de 3 aa básicos (Glu – Cys
– Gly), al ver su biosíntesis tiene un precursor q es el glutatión, lo
q la fitoquelatina toma del glutatión son las moléculas de Glu y Cys.
Atrapa los metales porque la fitoquelatina tiene grupos sulfhidrilos
con gran afinidad por los metales.
Análisis cinético de la absorción celular
(hoja 55 Fig. 3.5) Si representamos la velocidad de toma en función de
la concentración iónica nos da una cinética muy semejante a la
enzimática. (hoja 58 Fig. 6.21) Toma de sacarosa por hojas de soja.
Puede ocurrir q en el punto donde la velocidad se hace constante , esa
nos vuelva a aumentar. Esto es porque tenemos dos tipos de transporte,
uno el propio transporte y otro la difusión, de este modo podemos
diferenciar entre el transporte pasivo y el activo. (Fig. 6.22)
también podemos encontrar graficas mas complicadas, en esta primera
parte hay una cinética de saturación y luego se da otra fase. Esto nos
dice q hay dos transportadores, uno de baja afinidad y otro de gran
afinidad. El transportador mas efectivo dependerá de la concentración
q haya, a veces el segundo transportador solo se sintetiza cuando la
concentración iónica es muy alta.
Factores ambientales q afectan en la absorción.
- O2 : En presencia de O2 es un metabolismo aerobio donde hay ATP y
por tanto transporte y mayor absorción.
- CO2 : una planta en ausencia total de CO2 no fotosintetiza, gasta
todos los esqueletos carbonados, no se da la respiración no hay ATP y
no hay transporte activo q es el q mueve la mayor parte de los iones.
- Temperatura : la absorción aumenta con la Tª, pero hay un limite. A
partir de 40ºC esto cambia pues están implicadas proteínas y al
aumentar la Tª las membranas se hacen mas fluidas y los iones tienen
mas problemas para pasar.
- Luz : cuanta mas fotosíntesis mas ATP y por tanto mayor transporte.
Hay determinadas radiaciones PAR (radiación fotosintética activa) q
están en la zona en q vemos. Se aumentamos el PAR la fotosíntesis
aumenta, aumentando el ATP y el transporte aumentando la absorción. Si
aplicamos UV todo se destruye y disminuye la absorción.
- Inhibidores metabólicos : la absorción necesita ATP
mayoritariamente. El inhibidor afecta negativamente en la fotosíntesis
y por ello disminuye la absorción.
- Relación superficie –volumen de una célula : absorben mas q células
con menor relación. Una célula pequeña tiene mayor tasa de absorción.
- pH : altos pH impiden la absorción de aniones. Bajos pH impiden la
absorción de cationes. Si un ion K+ tiene q pasar la membrana va a
tener competencia con iones de su misma carga. El K+ para atravesar la
membrana tiene como competencia a altos pH a los protones y a bajos pH
a los hidroxilo.
- Presencia de otros iones : la presencia de iones similares dificulta
la absorción de elementos esenciales.
Absorción y transporte de iones en la planta
A excepción del C y un poco de O y de H la mayor parte de los
elementos la planta los tiene q tomar del suelo. Solo nos importa el
suelo en contacto con la planta. Presencia y disponibilidad de un
elemento no es lo mismo. El elemento tiene q estar y ser disponible
para poder pasar a la planta, tiene q ser soluble y pasar con el agua,
tiene q estar en forma iónica. (hoja 59 Fig. 11.4) De todo el suelo,
el importante para la planta es la Rizosfera (suelo controlado por la
actividad radical). Tanto la materia orgánica como la inorgánica del
suelo están cargadas negativamente. Esto tiene como consecuencia q a
las partículas del suelo se pegan cationes siendo adsorbidos y se
repelen los aniones. Los aniones han de estar en el agua q esta
disuelta en el suelo.
Capacidad de intercambio catiónico
Es la capacidad de las partículas del suelo para retener cationes. Los
aniones no suelen tener cargas + donde pegarse, por eso tienen q estar
flotando. /hoja 60 Fig. 5.3) En la raíz hay diferentes zonas. Hay una
zona fuera de la raíz cubierta por una especie de moco, es el mucigel,
son mucopolisacáridos secretados por el aparato de Golgi q se secretan
al exterior.
Los nutrientes con el agua pasan a favor de gradiente a la rizosfera.
Los carbohidratos sintetizados en la fotosíntesis llegan a emigrar por
la raíz. Parece q la planta exporta para el metabolismo de otros
organismos.
Funciones del mucigel
- Lubricación del ápice radical para favorecer su concentración.
- Evitar la desecación del ápice radical.
- Mantenimiento de una microflora (la mayor parte de las setas están
asociadas a árboles formando las micorrizas. Las Rhizobacterias
relacionadas con plantas q se aprovechan de los carbohidratos del
mucigel.
- Aumentar la disponibilidad de los nutrientes. Hay cambios de pH q
hacen disponibles nutrientes q no lo eran.
- Quelación de metales tóxicos. Estos polisacáridos son capaces de
fijar los metales e impedir q entren en la planta.
Toma de aniones y cationes
(hoja 59 Fig. 11.4) desde el punto de vista de la raíz los cationes
(+) están fijados y los aniones (-) no (estos dependen del estado
hídrico, cuando llueve los aniones son arrastrados perdiéndose
continuamente).
La mejor forma asimilable de N es el nitrato pero se dan problemas de
lixiviación (perdida del N con el agua gravitacional q se va
perdiendo), también esta el problema de la desnitrificación porque hay
bacterias q lo usan como aceptor de e- generando óxidos de N
eliminándose el nitrato del suelo.
El amonio es mas estable, pero al tener carga + puede ser volatilizado
y es un forma menos asimilable por la planta ya q es mas toxica.
La raíz para tomar los cationes tiene q romper el enlace por dos
procesos:
- Intercambio de contacto: la raíz suelta H+ para estabilizar las
cargas.
- La raíz es una fuente de CO2: el CO2 con el agua del suelo forma
H2CO3- H+ + H CO3- , fuente de protones q viajan al suelo y pueden
cambiarlo por el K+ pasando los K+ a la raíz mediante una disolución
diluida.
Distribución de los iones (hoja 61 fig5)
Una vez dentro los iones tiene q haber un movimiento radial para
llegar al xilema y luego un movimiento longitudinal hasta la parte
aérea.
Para viajar por el xilema hay dos rutas básicas:
- Vía del simplasto: es la vía citoplasmática a través de los
plasmodesmos, es poco efectiva y mas lenta q la del apoplasto. Esta
vía en algún momento puede tomar iones esenciales q necesita y repeler
no esenciales.
- Vía del apoplasto : Este ion en un momento dado puede salir al
exterior y viajar por las paredes sin entrar en ninguna célula, seria
por el xilema, sin atravesar los citoplasmas. Esta vía aumenta la
superficie de absorción, el problema es q no hay selectividad.
Hay una estructura especial q es la endodermis q obliga a los iones a
pasar por el simplasto pues de esta forma se da la selectividad y no
se cuelan iones no esenciales.
Para q esto se de, se tienen q sellar las paredes para q no entre el
agua, esto se hace con la suberina y la lignina, de manera q se obliga
al agua y a los iones a pasar por la membrana.
La endodermis tiene unas células selladas formando la Banda de Caspary
q hace a la pared impermeable al agua (apoplasto). Una vez atravesada
la endodermis pueden seguir por el simplasto o salir de las células y
seguir por la vía del apoplasto. La epidermis es uno de los pocos
puntos donde la pared esta sellada. La sabia del xilema es una
disolución muy diluida, tiene iones, nutrientes, ac. orgánicos
implicados en la translocación de algunos elementos, aa.
El nitrato es transformado en la raíz formando los aa q son los q
viajan por el xilema, no viaja el nitrato como tal. También viajan
hormonas (excepto el etileno).
La fuerza q dirige el movimiento hacia arriba es la transpiración. Una
vez en las hojas el agua se pierde, pero ¿q ocurre con los iones?.
Hay una serie de criterios q obligan a los iones a moverse hacia un
lado u otro. Se rige por la actividad metabólica q condiciona la
distribución iónica. Los tejidos jóvenes tiran de los iones porque
necesitan de ellos para formar sus estructuras. Los maduros como ya
están formados no tiran de estos iones ya q no tienen q hacer
estructuras nuevas. El superávit de iones se acumula en las vacuolas.
La deficiencia de iones móviles se nota primero en los tejidos
maduros.
Iones móviles : nitrato, potasio y fósforo.
Iones no móviles : azufre, hierro, manganeso y calcio.
Si la deficiencia se da en los tejidos jóvenes se trata de una falta
de iones no móviles.
Absorción foliar
Se da en todas las plantas, pero mas importante en las acuáticas y en
las epifitas aéreas. Tienen absorción foliar de todo tipo de
sustancias, ya sean orgánicas o inorgánicas. En ocasiones este tipo de
fertilización puede ser más favorable ya q en el suelo puede q haya
problemas para tomas los elementos.
Entran por tres vías :
- Estomas : si pulverizamos un pesticida algunas gotas caen sobre
ellos penetrando.
- Ectodesmos : discontinuidades de la epidermis por las q entran estas
gotas (vía especifica).
- Cutícula : es difícil, la translocación es lenta. Para q sea mas
rápida se le puede colocar al herbicida un compuesto apolar para q
pase la cutícula o se quede en ella, ya q la cutícula es apolar. Estas
moléculas son los surfactantes.
Nutrición mineral
La nutrición son los procesos metabólicos q favorecen la toma de
nutrientes a la planta y la incorporación a su interior. El suelo les
sirve como fuente de agua , nutrientes, sostén, gases. Mas de la mitad
de la superficie radical se debe a los pelos radicales, las raíces
leñosas no tienen importancia en cuanto a la absorción.
Disponibilidad de nutrientes
La disponibilidad de un elemento se debe a una serie de factores:
- pH : (hoja 62 Fig. 5.6) no todos los elementos se necesitan en igual
proporción, hay macro y microelementos. La disponibilidad se muestra
por la anchura de la banda. De 5 – 7 podrían ser los pH relativamente
buenos para el crecimiento del vegetal. Del pH depende la
disponibilidad, pudiendo inducir una deficiencia o toxicidad. Así para
tratamientos de suelos con metales pesados se sube el pH del suelo
para evitar la disponibilidad del mismo.
- Presencia o no de microorganismos en el suelo : los M.O. afectan a
través del pH del suelo subiéndolo o bajándolo. A veces liberan
compuestos carbonados o sustancias ácidas en su actividad metabólica
(terapia de suelos contaminados). Transformación de nutrientes. Una
planta cuando se muere se degrada dando una cantidad de nutrientes con
nitrógeno por la amonificación. Así una sustancia poco asimilable por
la planta se transforma en asimilable.
- Distribución de los elementos : alrededor de la Rizosfera se forma
una distribución libre de los elementos. El suelo depende de la
meteorización de las partículas q lo forman. Existen relaciones
simbióticas q pueden explorar a metros.
- Relaciones hídricas : permiten la movilidad de los iones,
relacionado con la distribución de los elementos. Cuanto mayor sea el
Ψ mas solubilidad y mayor facilidad del paso de cationes a la planta.
- Presencia de iones similares : la disponibilidad de algunos
elementos depende de la presencia de otros.
Determinación de un elemento esencial o no : si esta en el interior de
la planta, esta disponible y puede ser esencial para la misma. Son
necesarios unos requisitos para saber si es esencial (hoja 62).
• Favorece el crecimiento de la planta. Al variar la concentración del
nutriente crecerá hasta un tope q será la dosis optima y la
concentración critica q es la concentración de un elemento q con su
mínima concentración produce el máximo crecimiento.
• Micronutrientes son los q si los ponemos en exceso resultan tóxicos,
a pesar de ser esenciales. La grafica q harían es de este tipo:
El 96% de toda la materia de un vegetal lo constituye el oxigeno,
carbono e hidrógeno.
Elementos de esencialidad restringida. Solo son esenciales para un
grupo restringido de plantas : Na+ para las halófilas, Se para el
genero Astragalus , los animales q las comen se mueren por acumulación
de selenio.
Clasificación actual según su función (hoja 64 tabla 5.3)
- Grupo 1 : tomados en forma iónica o gas. Constituyentes mayoritarios
de la planta. Se incorporan por reacciones de carboxilación y
oxidorreducción.
- Grupo 2 : forman parte de reacciones de transferencia de energía.
- Grupo 3 : sin función conocida. Se toman en forma iónica. Relación
con el potencial osmótica.
- Grupo 4 : se asimilan en forma iónica o en quelatos. Son grupos
prostéticos implicados en la CTE.
Criterios de esencialidad de Arnon y Stout
Se han de cumplir tres postulados:
1. sin ese elemento la planta no completa su ciclo vital.
2. la deficiencia solo se corrige añadiendo el elemento.
3. se tiene q demostrar un papel directo nutricional en algún proceso
del vegetal.
Agentes quelantes (Fig. 5.10)
La forma iónica a veces es un problema. Un agente quelante es un
agente q se combina con el elemento. Uno de ellos es el EDTA
(etilendiaminotetraacetato). Es un compuesto carbonado cuyos grupos
carbonados se unen al Fe3+ haciendo el complejo muy soluble en agua
para q la planta lo adquiera. Se dan dos tipos de estrategias:
• Tipo 1 : en dicotiledóneas y monocotiledóneas. Se da cuando el pH es
alto. A través de la Rizosfera segregan ácidos orgánicos (cafeico), q
se combina con el Fe3+ formando el complejo cafeico Fe3+ . Este va por
la disolución cerca de la raíz y con agentes reductores donando e-
rompen el enlace liberando el cafeico y el hierro es absorbido.
• Tipo 2 : en gramíneas. La planta sintetiza compuestos llamados
fitosideróforos (ácido avénico y ac. mugineico), q se secretan a la
Rizosfera y se unen al Fe o a otro elemento. Pasa a la planta donde es
absorbido el complejo, se da la reducción y se asimila el compuesto
liberándose el ácido al suelo.
Funciones de los elementos (hoja 65, 66 y 67)
N : la deficiencia provoca clorosis. Reducción de crecimiento.
Esencial para aa, proteínas. Necrosis en las puntas. Coloración
purpúrea. Efectos detectados en las hojas maduras.
P : clorosis no tan evidente. Necrosis de puntas. Color verde intenso
con rojo.
K : clorosis marginal, seguida de necrosis marginal. Mantenimiento
osmótico, estabilización de membrana.
S : clorosis en las hojas mas jóvenes.
Ca2+ : la concentración esta regulada por la calmodulina. Los niveles
de Ca solubles son muy bajos. Implicado en la síntesis de 2º
mensajeros químicos. Necrosis en la zona interna de la hoja.
Mg2+ : tiene q ver con la activación de enzimas como la rubisco.
Clorosis q se nota en las hojas maduras y es de tipo intervenal.
Fe : se manifiesta en hojas jóvenes. Son la mayoría
de las clorosis q se ven. El 80% del Fe foliar esta en el cloroplasto
y son estos los q mas sufren la deficiencia. Cloroplastos mas
pequeños. Mas clorosis con baja cantidad de clorofilas y bajas tasas
fotosintéticas.
B+ : se ve en tejidos jóvenes. Aparecen necrosados y las plantas
tienen enanismo.
Mn2+ : en tejidos jóvenes clorosis con necrosis intervenla.
Cu2+ : plantas pequeñas y necrosadas en las puntas. Actúa como
cofactor en transporte de electrones (plastocianina).
Zn2+ : enanismo, acortamiento internodal.
Presencia de los elementos
La presencia de algunos elementos hace q la planta este mas
restringida.
• Toxicidad : metales pesados y salinidad. Q sean tóxicos no quiere
decir q no sean esenciales. Hay no esenciales q provocan toxicidad.
Metales pesados : hay q especificar si se trata de esenciales o no. Es
el tipo de contaminación mas grave. Como no se pueden degradar son
reconvertidos en compuestos mas utilizables por las plantas. Un
mecanismo de detoxificación es la fitorremediación. Los elementos mas
tóxicos son los q mas rápido se mueven para alterar puntos clave de la
planta (hoja 68 Fig. 8.8) dos estrategias para sobrevivir en un suelo
contaminado por metales pesados.
1. acumulación : se concentran en ciertos lugares gran cantidad pero
sin afectar a la planta. ( compartimentación en la vacuola o
precipitación con fosfatos impidiendo q afecte al metabolismo
Quelación y acumulación en las vacuolas)
2. exclusión : impedir q el metal este en el citosol. Otra técnica es
la fitovolatilización (Hg, Se). (unión de metales a la pared
transporte reducido a través de la membrana celular salida por
transporte activo de los metales).
Salinidad : hay q tener en cuenta el componente osmótico y el fenómeno
de la competencia. La presencia del ion baja el componente osmótico
porque restringe el agua. En cuanto a la competencia el Na+ esta
compitiendo con otros elementos esenciales.
Una estrategia para evitar la salinidad tiene q ver con la caída de
las hojas.
• Simbiosis : fijación biológica del N2 , micorrizas, líquenes,
parasitismo, carnivorismo.
Fijación biológica del N2 : asociación simbiótica entre hongos y
plantas. Ectomicorrizas y endomicorrizas. Las bacterias forman nódulos
en las raíces donde se fija el N2 , el amonio en aa, estos pasan a la
planta q cede carbohidratos a la bacteria.
Micorrizas : asociación simbiótica entre hongos y plantas. Pueden ser
Ectomicorrizas y endomicorrizas. (hoja 69) El nódulo tiene forma de
“Y” en las ectomicorrizas, y hay transferencia de los elementos. A la
planta entra agua y sustancia minerales y la planta exporta
carbohidratos. El hongo puede explorar suelos pobres en compuestos
orgánicos y la planta explora zonas lejanas con gran superficie de
exploración gracias a las hifas del hongo. El 90% de esta asociación
son endomicorrizas. Se puede dar simbiosis con mas de una especie de
hongos.
Ectomicorrizas : (hoja 70) el manto son las hifas del hongo q
envuelven a la planta. La red de Harlig es una red constituida por las
hifas del hongo q se introducen dentro por lugares determinados
(espacios intercelulares).
Endomicorrizas o vesiculo-arbusculares : (hoja 71) las hifas se
introducen en el interior. Las hifas van por los espacios
intercelulares dando la asociación mas intima. Degradan la pared y
obligan a la planta a invaginar su membrana pero nunca llegan al
citosol. El intercambio es mayor, siendo mas eficientes. No se sabe
bien como se da la transferencia, la planta degrada el arbúsculo y
absorbe los nutrientes. La mayor parte de las herbáceas hacen esto.
(hoja 72) La planta da sacarosa y el hongo fosfato. La sacarosa se
mueve a favor de gradiente expulsándose al exterior porque el hongo lo
toma desde el apoplasto mediante transporte activo. El fosfato desde
el hongo va por transporte pasivo gracias a una ATPasa q crea un
gradiente de fósforo entrando los fosfatos en la planta por simporte.
Líquenes : (hoja 73 Fig. 25.8) asociación entre un hogo y un alga. En
la matriz del hongo se insertan las algas. Hay una capa q le permite
unirse al sustrato, una capa de algas y luego una capa libre de algas.
El alga sintetiza esqueletos carbonados q difunden al hongo donde los
usa generando CO2 por la respiración y en vez de ir a la atmósfera lo
toma el alga volviendo a generar los esqueletos.
Parasitismo : hay especies q tienen un retroceso evolutivo en sus
procesos nutricionales, convirtiéndose en plantas parásitas ya q no
pueden tomar los elementos del suelo. Dos tipos de parasitismo:
Hemiparasitismo : muérdago. Parasita a nivel de xilema. Si tiene q
fotosintetizar. Fotosintetiza cuando otras plantas descansan. Su
estrategia es bajar mucho su ψ y tener abiertos los estomas mucho
tiempo transpirando a tasas muy elevadas (ψ muérdago < ψ planta)
Holoparasitismo : es la mas estricta. Parasita el floema. Ha perdido
las hojas pues no le hace falta fotosintetizar. El haustorio es una
prolongación q emite la holoparásita q le sirve para explorar el
medio. Lo único q sobresale es la flor. En el haustorio se consigue q
el ψ sea mas bajo y q las tasas de transpiración sean mucho mas altas
(mas q las tasas de transpiración de las hojas). Con las altas tasas
de transpiración la Tª de las hojas del parásito es mucho mas baja.
Carnivorismo : plantas q viven en turberas. Son zonas pobres en N.
Cazan insectos como fuente de nutrientes, sobre todo de N. Han
desarrollado enzimas para poder degradar los aa y obtener el nitrato.
Transporte por el floema
Estructura, composición, carga y descarta, funcionamiento.
El floema es un medio de transporte a larga distancia. Tiene un
movimiento ascendente, descendente y lateral. Necesita transporte
activo para bombear los solutos. Se mueve de las fuentes a los
sumideros.
Estructura
Localización mas extensa q el xilema
. Su unidad funcional es el elemento criboso q son células q se
adaptan para funcionar en este sistema, el elemento criboso tiene las
células acompañantes, las fibras o las esclereidas. La placa cribosa
conecta dos elementos cribosos. El transporte se da con la acción
conjunta del elemento criboso y la célula acompañante. El soporte lo
dan las fibras y las esclereidas.
Para demostrar q el floema es un tejido transportador dos
experimentos:
Pinchando la planta se ve q sale liquido q es lo q transporta.
La técnica del anillado es quitar la corteza selectivamente, rompiendo
el tejido floemático y preservando el xilema. Al cabo de los años el
tejido se deformaba. Supusieron q la raíz vivía de algo q iba de
arriba abajo (hoja 74 Fig. 4.1) se daba un abultamiento en la parte
superior de donde se da el corte. Por lo q se sabe q había algo q se
transportaba de las hojas a la parte basal y es el floema.
Elemento criboso : varios unidos forman el tubo criboso. Las áreas
cribosa laterales son para interaccionar con las células acompañantes
(hoja 75 Fig. 7.5)
Según madura el elemento va perdiendo el contenido del interior. El
elemento criboso tiene membrana y pierde casi todos los orgánulos en
cambio las células acompañantes tienen todo tipo de orgánulos (Fig.
7.6).
Ontogenia (hoja 76 Fig. 4.2) la célula meristemática se divide
longitudinalmente. La edad de ambas células es la misma. Se dan unos
cambios. Un elemento criboso maduro tiene un engrosamiento de
membrana. Hay una gran deposición de material de la pared debido al
Ap. Golgi. No hay lignificación. Los poros provienen de antiguos
plasmodesmos q darán la placa cribosa. La calosa permite tapar heridas
y parar el transporte a larga distancia. Todos los orgánulos se van
perdiendo. Los elementos cribosos van acumulando una proteína P. El
núcleo va desapareciendo. El Ap. Golgi, RE... inicialmente son activos
para sintetizar material de la pared y en las etapas finales pierden
su actividad, degeneran y se pierden.
Aunque son células sin orgánulos son vivas y activas porque no han
perdido la compartimentación aportada por la presencia de la membrana.
Composición de la sabia floemática
Cualquier pequeño cambio da artefactos microscópicos, por lo q se
congela y luego se da el corte. La presión del floema es positiva por
lo q los pulgones solo tienen q pinchar y el floema fluye hacia ellos.
Para recoger la sabia se coge el pulgón y se anestesia con CO2
cortándose el cuerpo y dejando el estilete (hoja 77 Fig. 7.12). la
sabia del floema es mas concentrada q la del xilema.
De toda la materia seca la mayoría de lo q se transporta son
carbohidratos (disacáridos como la sacarosa) oligosacáridos como la
rafinosa, estaquinosa, verbascosa... también se transportan aa, los
mas abundantes son ac. glutámico, asp, asparagina. Son la fuente
principal de transporte de N. Hay gran cantidad de iones (nitrato a
través del xilema), las formas nitrogenadas viajan siempre en forma de
aa o proteínas. pH ligeramente alcalino. Transporta todas las hormonas
menos el etileno. Potencial osmótico bajo. (hoja 79 Fig. 6.12) las C4
son mas eficientes en el transporte q las C3. El transporte en cada
tubo es unidireccional, en algunos caso se puede dar una mezcla en las
zonas donde los tubos cribosos se comunican.
Carga y descarga
Puede ir en cualquier dirección. Siempre se mueve de la fuente al
sumidero. La fuente es el órgano productor o almacenador de
fotosintetizados (hoja metabolicamente activa). El sumidero es el
órgano importador de fotosintetizados (una hoja en formación). Una
fuente puede convertirse en un sumidero y un sumidero puede
convertirse en una fuente. (Fig. 6.11) el fruto es el sumidero. Una
patata en formación es un sumidero y cuando se diferencia es una
fuente. (Hoja 80) la carga se da de la fuente al tubo criboso. Ocurre
por:
• vía simplasto : célula del mesofilo célula acompañante tubo criboso.
• vía apoplasto: salen de la célula productora al exterior celular y
del apoplasto pasan al tubo criboso.
Carga : se genera transporte primario por la actividad de un ATPasa q
bombea P+ al exterior. Se cree q el mesófilo también contribuye con
otra ATPasa menos efectiva. Con esto el mesófilo tiene un pH alcalino
el del simplasto es un pH ácido. Los H+ tienden a entrar y al entrar
mueven un transportador q mueve la sacarosa.
Por este transporte también se pueden introducir aa en contra de
gradiente gracias al gradiente de H+. La sacarosa sale de la célula
productora. Esta se esta formando continuamente estando muy
concentrada en la célula generándose un gradiente por lo q la sacarosa
sale por un mecanismo de difusión facilitada sin gasto de ATP.
El gradiente se mantiene porque la sacarosa esta siendo retirada
continuamente del apoplasto hacia el tubo criboso.
Descarga : (hoja 81) bien por la vía simplástica o apoplástica. Un
sumidero consultivo es el q consume. Se da la vía del simplasto. Un
sumidero de reserva es el q acumula. Se da la vía del apoplasto.
En el simplasto la sacarosa puede salir como tal e incorporarse,
(tiende a salir del tubo criboso a favor de gradiente porque se esta
retirando continuamente). Se gasta cuando se carga, no cuando se
descarta. No se modifica en plantas C3.
Las C4 tienen un proceso más acelerado para crear el gradiente. En el
apoplasto se rompe la sacarosa en sus monómeros, entran en la célula
de almacenamiento y se vuelve a formar la sacarosa. Se crea un segundo
gradiente, en el mismo apoplasto se da el tirón para crear el
gradiente.
Funcionamiento
Teoría de flujo a presión o de flujo en masa. Hipótesis de Munch (hoja
82 Fig. 6.21) siempre se mueve de la fuente al sumidero. Primero hay
una carga y posteriormente una descarga. Se da la carga del floema.
Hay un sistema y concentra sacarosa en el tubo criboso disminuyendo el
Ψs. La gran concentración de sacarosa tira del agua de las células de
alrededor. Es una entrada pasiva de agua. Con la entrada de agua
aumenta el volumen distorsionando las paredes. Al no romperse las
paredes dado q están reforzadas el aumento de volumen acaba moviendo
en masa el floema dándose el transporte.
En el sumidero da sacarosa tiende a salir por si misma a favor de
gradiente. Hay un aumento del Ψs y hay una salida pasiva de agua a
favor del gradiente de Ψ q se genera.
Las paredes están relajadas y no crean ninguna fuerza. En cuanto se
empieza a consumir sacarosa el movimiento se invierte. El floema tiene
q tener unas observaciones q cumplan la hipótesis:
- Presión positiva (0.1 – 4).
- Los solutos tienen q viajar en el mismo sentido y velocidad.
- Tiene q existir un gradiente de concentración. Más concentrado en la
fuente. Menos concentrado en el sumidero.
- Poros abiertos. Las plantas no han quitado las placas cribosas
porque les permiten tapar fisiológicamente al ocluir los poros con
proteína P.
Otras teorías son:
Flujo electroosmótico : con la célula acompañante se genera un flujo
de K+ q por gradiente pasa al otro lado de la planta generando una
especie de gradiente eléctrico. Se genera un gradiente eléctrico q
permite el paso de sacarosa, aa, ...
Corrientes protoplasmáticas : en la célula se generaría algún
movimiento q permitiría mover la sacarosa en una dirección y por
ejemplo aa en dirección contraria. Esto explicaba la
bidireccionalidad.
Aspectos generales del metabolismo secundario
El metabolismo secundario es todo lo q no es metabolismo primario
(procesos básicos q tienen todos los organismos vivos). Los
metabolitos primarios por excelencia son lípidos, azúcares, proteínas
y ac. nucleicos. Un metabolito 2º es uno q deriva de uno 1º o de una
ruta colateral. (los 2º son tan importantes como los primarios.
Características de los metabolitos secundarios
Origen mayoritariamente vegetal. Existen pocos microbianos e
inexistentes en animales.
Estructura muy heterogénea. Hay compuestos muy sencillos y otros muy
complejos. (hoja 83)
Distribución heterogénea entre las especies y también dentro de una
especie en los diferentes tejidos de la planta. Esto no ocurre con los
metabolitos primarios.
Están compartimentizados. Los 2º están en un lugar encerrados por
membrana porque son tóxicos. La mayor parte de ellos están en la
vacuola. Un carotenoide esta en el cloroplasto, unos implicados en la
fotosíntesis y otros en dar color.
Les permite regular su metabolismo: enzimas en el citoplasma inactivo
porque el sustrato esta en un orgánulo.
La producción se produce en células especializadas y no todas las
células están especializadas en su producción. Su producción además
depende de la fenología, esto es, de los diferentes estadios del
desarrollo de la planta. Su producción depende del ambiente. Suele ser
un tipo de respuesta ante un estrés. A veces el ambiente puede ser
otra planta, un patógeno o incluso un animal.
Tipos (hoja 84)
Fenoles : tienen un anillo bencénico.
Terpenoides : β-caroteno esta en todos, es el centro de reacción del
fotosistema II.
Alcaloides : tienen nitrógeno en las moléculas.
aa no proteicos.
Glucósidos cianógenos : compuestos con actividad ciano.
(hoja 85 Fig. 13.4) Algunos metabolitos 1ºs se desvían hacia otras
rutas dando lugar a compuestos que son los metabolitos 2ºs y también
algunos metabolitos 1ºs como son los aa aromáticos y los alifáticos se
generan los alcaloides.
Funciones de los metabolitos secundarios
Su primera función es q no tengan ninguna función especifica. Hablamos
de:
Funciones itraespecificas.
- Desintoxicación de metabolitos 1ºs: estos compuestos son importantes
dentro del vegetal.
Las plantas no tienen sistema excretos. En ocasiones aprovechan la
abscisión foliar para expulsar elementos tóxicos. Otras veces se
desembarazan de elementos tóxicos provenientes del metabolismo 1º.
La ruta del ácido Shikimico nos da aa aromáticos como Phe, Tyr, y Trp.
Si hay una proteolisis rápida se liberan aa q pueden ser tóxicos. Una
forma de eliminarlos es desaminarlos y dar lugar a fenoles, esto es
transformarlos en otros compuestos.
En condiciones de sequía la [fenoles] es mayor. Con el amonio q se
libera podemos incluirlo y formar alcaloides.
Lo q pueden hacer es compartimentalizarlos y acumularlos por ejemplo
en el exterior de la célula como la lignina .
- Importantes en la fisiología celular : en la fotosíntesis hay
compuestos necesarios para q se pueda dar. Otro ejemplo son las
vitaminas.
- Importantes en la estructura : algunos compuestos son importantes
para la estabilización de las membranas, lignina, celulosa.
Funciones intersepecificas.
Importantes para controlar a otras especies. Importantes en su
relación con otras especies.
Hoy unas teorías de Bell (1978) q proponía q la distribución era
debida a las funciones interespecificas.
Todo este desarrollo de la presión selectiva se ve representada por la
inmovilidad de las plantas.
En otros organismos la movilidad les permite defenderse. las plantas
han de defenderse valiéndose de la movilidad de otros organismos. Las
plantas aprovechan la movilidad de otros organismos para poder
transportar sus genes. Hay una coevolucion.
La hoja se transforma en flor, el insecto se acerca a recoger el polen
y transmite los genes de planta en planta.
En el caso de la defensa ocurre lo mismo. Formaría compuestos muy
tóxicos q impidan ser depredados por otros organismos.
Otro ejemplo es la defensa contra organismos patógenos. Es el caso de
las fitoalexinas, esto es, producen fitoalexinas q les sirven como
mecanismo de defensa.
Las plantas viven en competencia constante bien mediante una guerra
con las mismas armas o bien porque han desarrollado unas rutas q les
permiten obtener compuestos q actúen en la fisiología de otras
plantas. Es el caso de las fitolexinas q son compuestos con actividad
antimicrobiana q pueden no ser tóxicos para ella misma.
Puede darse la producción de compuestos q afecten a otras plantas q
impiden de un modo restringido su crecimiento. Son compuestos
aleloquimicos.
La alelopatia es la ciencia q estudia el efecto de agentes químicos de
distintas plantas sobre el desarrollo de otras plantas.
La juglona es uno de estos agentes aleloquimicos. El ac. cinámico, ac.
formica, ac. felúrico, en definitiva ácidos benzoicos son agentes
aleloquimicos.
Una vez q las hojas del nogal caen siguen funcionando para el árbol.
La degradación de estas hojas impide la germinación de semillas de
otras plantas.
Las plantas son capaces de emitir compuestos q son los flavonoides q
actúan en la simbiosis.
También en una labor previa a la síntesis de flavonoides facilita el
crecimiento de alguna especie q va a ser el q intervenga en la
simbiosis.
La planta sintetiza compuestos q atraen a organismos. También se da la
presencia de compuestos repelectantes para q no sean depredadas.
Compuestos Fenolicos
Presencia de un anillo aromático con una serie de radicales. Su
localización en la planta es mayoritariamente vacuolar. En un
metabolito 2º la parte no azucarada de la molécula, es la aglicona q
esta aislada y además activa.
Son moléculas muy reactivas ya q son capaces de establecer muchos
puentes de H. Rompen la estructura tridimensional de las proteínas
desnaturalizándolas.
Dada la presencia de tantos OH e H son capaces de quelar metales
quedando desnaturalizados muchos enzimas q actúan con metales.
Presentes en organismos vegetales y no en animales.
Biosíntesis de fenoles
Siempre provienen de esqueletos carbonados.
Provienen de la ruta del ac. Shikimico. Este ácido esta en todas las
plantas. Se descubrió en la planta Ilicium amisalium donde su
concentración es muy alta.
Proviene de intermediarios de algunas rutas del metabolismo primario
principalmente del ciclo de Calvin y glucolisis q originan la
eritrosa-4p y el PEP respectivamente.
La concentración de la eritrosa-4p y PEP da una cadena corbonada de 7
C.
Mediante transformaciones se origina el shikimato q sufra
fosforilación dando shikimato 7P q con otro PEP da
enolpiruvilshikimato-P. A partir de este se origina el corismato.
Dependiendo del enzima q coja de este corismato dará un compuesto u
otro. Si lo coge la antramilato sintasa da Trp y di lo coge la
corismato sintasa dará o bien Tyr o Phe.
Ya tenemos los aa aromáticos esenciales.
El Trp se utiliza para la síntesis de AIA, proteínas y alcaloides, con
la otra ruta damos lugar al resto de las enzimas.
Esta parte del ciclo se abre con el enzima Pal (Phe amonioliasa) q
desamina a la Phe y se convierte en un fenol q es el cumarato.
Si utiliza Tyr la desamina y da cinamato.
Con estos fenoles hacemos lignina, muchos alcaloides y muchos
flavonoides.
(hoja 87) Dependiendo de los radicales q tenga y del enzima q actué
obtendremos el resto de los fenoles. Los fenoles simples son los más
tóxicos.
Funciones
- Pigmentos florales : los más aparentes son las antocianinas q es una
antocianina con un azúcar. Unas dan color rojo, otras azul, pueden
cambiar de color dependiendo del entorno fisiológico de la planta sin
cambiar de moléculas. Otros fenoles son flavonoles y flavononas.
- Pigmentos de los frutos : antocianinas. Se ven porque la clorofila
se va degradando. También están las isoflavonas.
- Reconocimientos de simbiosis : flavonoides. Agentes alelopaticos.
- Protectores contra herbívoros e insectos : flavonoles.
- Agentes funguicidas : isoflavonas.
- Fitoalexinas : isoflavonas.
- Estrés abióticos : Luz , frió., inducen la formación de fenoles.
Compuestos Isoprenoides O Compuestos Terpenoides
Grupo con mayor numero de representantes conocidos.
Insolubles en agua más q los fenoles.
Estructuras muy variadas de unos compuestos a otros.
Origen biosintético q es la ruta del acetato-mevalonato.
Biosíntesis
Si son 5 o múltiplos de 5 el numero de C es un terpenoide ya q todos
son repeticiones de una unidad de 5C q es el isopreno.
Todos los compuestos son repeticiones de esta estructura.
Ruta clásica
Se condensan 3 moléculas de Ac CoA dando una molécula de 6C (ac.
Mevalonico). Este se fosforila 3 veces y se descarboxila
transformándose en una molécula de 5C q es el isopentenil PPi. Este se
parece al isopreno y es la molécula q se va repitiendo. este
isopentenil PPi es el intermediario a partir del cual se forman los
demás.
Ruta alternativa
Se forman 3 P-gliceratos a partir de la glucólisis. También se
necesita piruvato, pero solo se necesitan 2C de este pirúvico. La
unión da un compuesto de 5C q a el deoxixilulosa 5P y a partir de esta
el isopentenil PPi. Este intermediario es común a ambas rutas.
(hoja 88)biosíntesis de terpenoides: el fitoeno es el primer
intermediario tetraterpeno. El diterpeno se forma uniendo dos
moléculas de geranil PPi. Solo se acumula el producto final, no los
intermediarios. Toda esta síntesis se da en el plastidio.
El enzima, sin soltar al monoterpeno, le une un IPP originando un
diterpeno q es el q suelta.
En la ruta clásica los sesquiterpenos se liberan. En el plastidio los
hemiterpenos y monoterpenos se pueden liberar.
Los hemiterpenos son muy volátiles. Los sesquiterpenos no se pueden
liberar, esto es al unirle al monoterpeno un IPP este no se libera,
pero al unirle otro IPP formamos los diterpenos q sí se pueden
liberar.
Tipos y funciones
Monoterpenos
Son moléculas de 10C. Forman la mayor parte de los aceites esenciales.
Le sirven como:
- Alelopaticas
- Insecticida
- Antialimentario
- Atrayentes
- Repelentes
- Bactericidas
- Funguicida
Sesquiterpenos
- Fitol y ac. giberelico
- Funciones en la fotosíntesis , síntesis de la clorofila.
- Antialimentario (resina)
- Antioxidantes
- Antimitotixos
Triterpenos
- Insecticidas
- Antialimentario
Tetraterpenos
- Pigmentos accesorios
- Centro de reacción en la fotosíntesis
- Antioxidante
ESPAI RESERVAT PER L’ÍNDEX
ESPAI RESERVAT PER LA TAULA
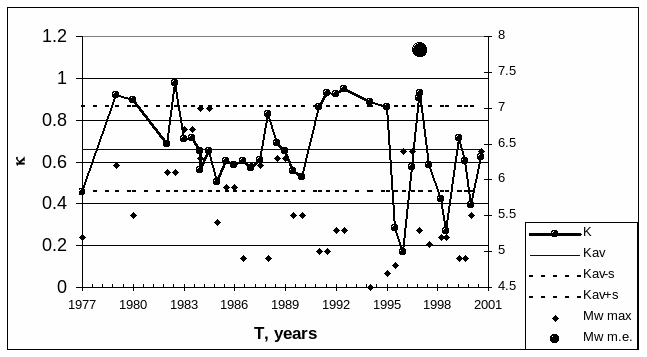 SEISMIC MONITORING OF STRONG EARTHQUAKES SOURCE AREAS A LUTIKOV
SEISMIC MONITORING OF STRONG EARTHQUAKES SOURCE AREAS A LUTIKOV PROJETO EDI TRANSPORTE OCOREN – OCORRÊNCIA NA ENTREGA DE
PROJETO EDI TRANSPORTE OCOREN – OCORRÊNCIA NA ENTREGA DE STRUCTURE AND BONDING 1 ONLY ONE OF THE FOLLOWING
STRUCTURE AND BONDING 1 ONLY ONE OF THE FOLLOWING UNITED NATIONS ECETRANSWP29GRE201217 ECONOMIC AND SOCIAL COUNCIL DISTR GENERAL
UNITED NATIONS ECETRANSWP29GRE201217 ECONOMIC AND SOCIAL COUNCIL DISTR GENERAL PLA OPERATIU DE LA BIBLIOTECA DE CATALUNYA 2012 OBJECTIU
PLA OPERATIU DE LA BIBLIOTECA DE CATALUNYA 2012 OBJECTIU MENGOPERASIKAN SOFTWARE SPREADSHEET (MS EXCEL 2007) DISAJIKAN UNTUK MEMBANTU
MENGOPERASIKAN SOFTWARE SPREADSHEET (MS EXCEL 2007) DISAJIKAN UNTUK MEMBANTU RESPUESTAS EJERCICIO Nº 2 – OPOSICIONES AÑO 2000 1
RESPUESTAS EJERCICIO Nº 2 – OPOSICIONES AÑO 2000 1 FACULTAD DE MEDICINA HUMANA ESCUELA PROFESIONAL DE MEDICINA HUMANA
FACULTAD DE MEDICINA HUMANA ESCUELA PROFESIONAL DE MEDICINA HUMANA UNIT R087 – CREATING INTERACTIVE MULTIMEDIA PRODUCTS CREATING
UNIT R087 – CREATING INTERACTIVE MULTIMEDIA PRODUCTS CREATING CHAPTER 4 A GRAPHIC REPRESENTATION FROM OUTER SPACE
CHAPTER 4 A GRAPHIC REPRESENTATION FROM OUTER SPACE VOLCARUNCOM UN EXEMPLE DE SORTIE AU VOLCAN HUIT ARRÊTS
VOLCARUNCOM UN EXEMPLE DE SORTIE AU VOLCAN HUIT ARRÊTS OBRA PINTURA INTERIOR Y EXTERIOR DEL TORNAVIAS PLIEGO DE
OBRA PINTURA INTERIOR Y EXTERIOR DEL TORNAVIAS PLIEGO DE L ESSON 8 HEBREWS 10131 PRAY OBSERVE INTERPRET APPLY
L ESSON 8 HEBREWS 10131 PRAY OBSERVE INTERPRET APPLY